
Анатомия и физиология млекопитающих

Анато́мия и физиоло́гия млекопита́ющих — совокупность сведений о строении и функционировании организма у млекопитающих.
Млекопитающие представляют собой один из классов позвоночных животных. С другими таксонами позвоночных их объединяет общность морфологической и физиологической организации. Для анатомии и физиологии млекопитающих характерно наличие тех же функциональных систем, что и у других позвоночных; однако многие из этих систем достигли у млекопитающих очень высокого уровня развития, и в целом данный класс считают наиболее высокоорганизованным по сравнению с другими классами позвоночных[1].
К основным отличительным морфофизиологическим особенностям, характерным для всех млекопитающих или для подавляющего большинства их представителей, относят: наличие молочных желёз и вскармливание потомства молоком, живорождение, истинную гомойотермию (теплокровность), наличие (хотя бы в эмбриональном периоде развития) волосяного покрова, высокую степень развития неопаллиума, составляющего у млекопитающих основную часть коры головного мозга, и ряд других особенностей[2].
Покровная система
Кожный покров
Кожа образует эффективный барьер между организмом и внешней средой, защищая его от нежелательных физико-химических воздействий, проникновения патогенов и от нерегулируемых потерь воды и солей[3]. Для кожи млекопитающих характерны значительная толщина, богатство роговых образований и наличие большого количества кожных желёз, весьма немногочисленных у пресмыкающихся и птиц. Как и у других позвоночных, она состоит из двух слоёв: верхнего эктодермального (эпидермис) и нижнего мезодермального (дерма, или кориум)[4].

- A — эпидермис;
- B — дерма;
- С — гиподерма;
- D — кровеносные и лимфатические сосуды;
- E — зародышевый слой.
- 1 — волос;
- 2 — роговой слой;
- 3 — пигментный слой;
- 4 — средний слой;
- 5 — базальный слой;
- 6 — выпрямляющая мышца волоса;
- 7 — сальная железа;
- 8 — волосяной фолликул;
- 9 — волосяной сосочек;
- 10 — нервное волокно;
- 11 — потовая железа;
- 12 — тельце Пачини;
- 13 — артерия;
- 14 — вена;
- 15 — чувствительное нервное окончание;
- 16 — папиллярный слой;
- 17 — пора потовой железы.
Эпидермис многослоен: в его основании расположен мальпигиев слой из интенсивно размножающихся цилиндрических или призматических эпителиальных клеток — кератиноцитов. Кератиноциты последовательно сдвигаются вверх, уплощаясь и подвергаясь всё большему ороговению, то есть заполнению кератиновыми волокнами (на эти волокна и связывающий их филаггрин приходится 80—90 % общей массы белка эпидермиса млекопитающих). На завершающем этапе ороговения кератиноциты теряют ядра, а мёртвые ороговевшие клетки постепенно слущиваются с поверхности кожи[3][5]. Окраску кожи млекопитающих обеспечивают меланины — пигменты чёрного, коричневого, ржаво-красного или жёлтого цвета; эти пигменты вырабатываются специализированными клетками мальпигиева слоя — меланоцитами — и накапливаются в особых органеллах — меланосомах, мигрирующих из меланоцитов в соседние кератиноциты[6].
В нижнюю поверхность эпидермиса вдаются сосочки дермы. Основную толщу последней образует сетчатый слой из перекрещивающихся коллагеновых и эластиновых волокон, он придаёт коже надлежащую прочность. Ниже сетчатого слоя лежит рыхлый слой соединительной ткани — подкожная жировая клетчатка, состоящая из жировых клеток (адипоцитов) и окружающих их волокон. Дерма пронизана кровеносными сосудами и нервными окончаниями; она служит защитой тела от механических повреждений[7].
Для млекопитающих весьма характерны роговые образования эпидермального происхождения: волосы, когти, ногти, копыта, рога и чешуйки[8].
Волосяной покров

Волосы — образование, присущее исключительно млекопитающим и, по-видимому, наиболее продвинутым терапсидам и не гомологичное каким-либо производным кожи других амниот. Их главная функция — теплоизоляция, хотя толчком для появления волосяного покрова послужила, скорее всего, осязательная функция[9].
Помимо термоизолирующей и осязательной функций, волосы защищают кожу от повреждений и паразитов, улучшают аэро- и гидродинамические свойства тела, обеспечивают видоспецифичность окраски. Отсутствие волос у ряда млекопитающих (слоны, сирены, часть носорогов, гиппопотамы, китообразные) носит вторичный характер[10]. Это можно сказать и о человеке, который после перехода от жизни в тенистых лесах к обитанию на открытых пространствах утратил бо́льшую часть волосяного покрова[11].
У большинства млекопитающих мех состоит из двух ярусов: верхний (шерсть) образуют длинные остевые волосы, нижний (подшёрсток) — более короткие и тонкие, волнообразно изогнутые пуховые волосы. У взрослых оленей, кабанов и настоящих тюленей летний мех образован одной остью, а у землероев (слепыши, кроты) мех состоит из одного пуха. Видоизменениями остевых волос являются щетина (свиньи, тенреки), иглы (ехидны, ежи, дикобразы) и вибриссы — длинные жёсткие волосы на конце морды, брюхе или конечностях многих зверей, служащие для осязания[12].
Окраска шерсти млекопитающих, как правило, является покровительственной. У некоторых представителей наблюдается сезонная смена окраски. Иногда окраска участвует в формировании вторичных половых признаков[13].
Волосяной покров периодически меняется. Смена меха (линька) может наблюдаться два раза в году: весной и осенью. Так происходит у белки, лисицы, песца, крота. У других, например сусликов, линька происходит раз в год — весной[13].
Другие роговые образования

У большинства млекопитающих концевые фаланги пальцев защищены когтями, которые остры и сильно изогнуты у древесных форм и хищников, уплощены и расширены у роющих форм[14]. У многих приматов когти преобразовались в плоские ногти, прикрывающие фалангу лишь сверху; у копытных в результате усложнения когтей сформировались копыта — толстые образования, которые играют роль своеобразного футляра для концевой фаланги, особо важный при беге и прыжках по твёрдому грунту[8].
У многих млекопитающих на хвосте (бобр, мышиные, шипохвостые, выхухоль, многие сумчатые) или конечностях (роющие формы) имеются роговые чешуйки. У панголинов и броненосцев крупные роговые чешуи покрывают всё тело, причём у броненосцев они подстилаются ещё костными щитками, являющимися производными дермы и образующими панцирь[8]. Чешуя млекопитающих вполне гомологична таковой у пресмыкающихся[13].
Для некоторых млекопитающих (носороги, вилороговые, полорогие) характерно наличие рога — массивного разрастания ороговевшего эпителия, используемого для защиты и нападения. Иное происхождение у ветвистых рогов оленей, которые развиваются из дермы и состоят из костного вещества; основное их применение — не оборонительное, а турнирное (используются в драках самцов за самку)[15][16]. Если у полорогих стержни рогов закладываются независимо от черепа и срастаются с ним позднее, то рога оленей развиваются как выросты лобных костей; при этом у большинства видов оленей каждую весну в результате резорбции костной ткани под действием гидролитических ферментов, выделяемых специальными клетками-остеокластами, происходит сбрасывание рогов, а затем они отрастают заново[17][18].
Цедильный аппарат китов — «китовый ус» — также образован роговыми утолщениями (производными кожных нёбных складок), которые превращаются в плоские и расщеплённые пластины, расположенные двумя рядами с каждой стороны ротовой полости. Наружный край этих пластин — жёсткий, а внутренний имеет густую бахрому, причём число пластин в каждом ряду может доходить до 430[19][20].
Экзокринная система
Экзокринную систему образуют железы внешней секреции, выделяющие вырабатываемый ими секрет через выводные протоки на поверхность тела или слизистых оболочек внутренних органов[21].
Сильное развитие разнообразных кожных желёз, образующихся из погружённых в толщу дермы эпидермальных зачатков, млекопитающие унаследовали через своих синапсидных предков от земноводных[22][23]. Эта черта отличает млекопитающих от других современных амниот, у которых кожные железы немногочисленны[24].
Различают несколько типов кожных желёз. Появление волосяного покрова привело к развитию сальных и потовых желёз, предназначенных для смазывания волос и контроля потерь тепла[22]. Протоки сальных желёз открываются в волосяные сумки; эти железы имеют гроздевидное строение и выделяют жирный секрет, который смазывает поверхность кожи и волосы, предохраняя их от смачивания и пересыхания, а также препятствует проникновению болезнетворных микроорганизмов. Протоки трубчатых потовых желёз открываются на поверхность кожи или в верхние части волосяных сумок; данные железы, имеющие трубчатую форму, секретируют пот, выполняя одновременно терморегуляционную и выделительную функции[25]. Под верхне-наружным краем глазницы располагаются слёзные железы альвеолярно-трубчатого строения, секрет которых предохраняет глаза от высыхания[26].

Млечные железы, представляющие собой видоизменённые потовые железы, в качестве секрета выделяют молоко — основную пищу детёнышей млекопитающих. Млечные железы сохранили трубчатое строение у однопроходных и приобрели гроздевидное строение у сумчатых и плацентарных. У представителей этих двух групп протоки млечных желёз открываются на сосках; млечные железы однопроходных сосков не имеют, и их протоки открываются в волосяные сумки, так что детёныши слизывают капельки молока с волос матери[27].
Соски имеются и у самок, и у самцов, у последних они рудиментарны и не способны выделять молоко. Исключение составляют два вида крылановых — бурый крылан (Dyacopterus spadiceus) и бисмаркская масковая летучая лисица (Pteropus capistratus): если у самки рождается двойня, то млечные железы самца начинают выделять молоко (хотя и в небольшом количестве), и он участвует в кормлении одного из детёнышей[28][29].
Количество сосков может варьировать у разных видов от 2 (высшие приматы, некоторые копытные и рукокрылые) до 24 (некоторые тенреки) и даже 27 (некоторые опоссумы), причём у сумчатых число и расположение сосков может быть непарным, а у плацентарных — всегда парное. У плодовитых форм сосков обычно много, и расположены они двумя рядами от передних конечностей до паха (насекомоядные, хищные, грызуны). Если же число сосков невелико, то они могут располагаться на груди (у высших приматов, сирен, слонов, многих летучих мышей), брюхе (у части низших приматов есть пара брюшных и пара грудных сосков) или в паху (у многих копытных, причём у большинства из них соски сливаются в единое вымя). Выводные протоки млечных желёз открываются на сосках либо отдельными отверстиями (у сумчатых, приматов, грызунов), либо в общий выводной канал (копытные, хищные)[30][31].
У некоторых млекопитающих (многих хищных, кабарги, бобров, ондатры, выхухоли и др.) имеются пахучие железы. Это — видоизменения потовых или сальных желёз или же комбинации тех и других. Они выделяют пахучие секреты, с помощью которых звери метят свою территорию, отличают особей своего вида, находят особь другого пола. Скунсы, мангусты, африканская ласка используют зловонный секрет таких желёз для отпугивания врагов[27][32].

Ядовитые железы, выделяющие достаточно сильный яд, найдены у утконоса, взрослые самцы которого с помощью расположенных на задних ногах ядовитых шпор убивают своих жертв. Среди ехидн такие шпоры встречаются и у самок, но яд, продуцируемый ядовитыми железами ехидн, намного слабее яда утконоса. Палеонтологи недавно обнаружили подобные шпоры у представителей различных групп ранних млекопитающих (докодонты, триконодонты, многобугорчатые, симметродонты), так что ядовитыми железами на ногах, скорее всего, обладал и общий предок класса Mammalia, однако сумчатые и плацентарные такие шпоры утратили[33].
Помимо этого, у млекопитающих имеется достаточно много и экзокринных желёз, отличных от кожных. В частности, функции желёз внешней секреции выполняют слюнные железы, печень и поджелудочная железа, секреты которых, попадая в пищеварительный тракт, играют важную роль в ходе пищеварения[34].
Скелет
Подобно другим позвоночным, млекопитающие имеют внутренний скелет, который выполняет функции опоры тела, субстрата для прикрепления мышц и защиты для ряда внутренних органов. У броненосцев есть также — как дополнение к внутреннему — и внешний скелет в виде панциря. По сравнению с рептилиями, у млекопитающих количество отдельных костных элементов в скелете сильно сокращено, а хрящ у взрослых особей сохраняется лишь в местах, требующих особой гибкости[35].
Череп

2 — лобная кость; 3 — слёзная кость; 4 — нёбная кость;
5 — скуловая кость; 6 — носовая кость; 7 — резцовая кость;
8 — теменная кость; 9 — межтеменная кость; 10 — чешуйчатая кость;
11 — затылочная кость; 12 — нижняя челюсть.
Череп млекопитающих имеет относительно крупную мозговую коробку, уже у позднетриасового морганукодона она в 3—4 раза крупнее, чем у терапсид того же размера[36]. Число отдельных костей в черепе меньше, чем у других позвоночных, что объясняется срастанием ряда костей между собой. Так, срослись воедино основная, боковая и верхняя затылочная кости, образовав единую затылочную кость; ушные кости срослись в единую каменистую кость. Крыловидная кость срастается с основной клиновидной, глазоклиновидная — с передней клиновидной. Тем не менее, у млекопитающих появляются и новые кости — решётчатая в носовой полости и межтеменная в крыше черепа[37].
Бока черепной коробки образованы чешуйчатыми костями; отходящие от них скуловые отростки соединены со скуловой костью, а та — со скуловым отростком верхнечелюстной кости; в результате образуется характерная для млекопитающих единственная скуловая дуга. Затылочная часть, как уже упоминалось, образована единой затылочной костью; для сочленения с первым шейным позвонком — атлантом — она снабжена двумя мыщелками (отметим, что пресмыкающиеся и птицы имеют один затылочный мыщелок[38]). Крышу черепа формируют парные теменные, лобные, слёзные и носовые кости, а также непарная межтеменная кость. Дно мозговой коробки составляют основная клиновидная и передняя клиновидная кость, дно лицевой (висцеральной) части образовано крыловидными, нёбными и верхнечелюстными костями. В дне черепа, рядом со слуховой капсулой располагается свойственная только млекопитающим парная барабанная кость, являющаяся гомологом угловой кости синапсид[37][39].
Уменьшается число отдельных костей и в челюстях млекопитающих: нижнюю челюсть образуют только две парные зубные кости, верхнюю — парные верхнечелюстная и предчелюстная. Нёбные отростки последних вместе с нёбными костями формируют вторичное костное нёбо, отделяющее ротовую полость от носовой и позволяющее не прерывать дыхание при пережёвывании пищи или сосании молока. Такое нёбо характерно именно для млекопитающих и их цинодонтных предков, но не для других амниот (хотя подобное нёбо независимо возникло у тероцефалов и некоторых пресмыкающихся, в том числе крокодилов, но у них оно устроено иначе)[40][41]. Соединение верхней челюсти с черепной коробкой — жёсткое (в то время как у клювоголовых, чешуйчатых и птиц она способна двигаться относительно других частей черепа), а нижней челюсти — подвижное, причём зубная кость причленяется непосредственно к чешуйчатой кости[42].
Сочленовная и квадратная кости, свойственные рептилиям и синапсидам, становятся элементами среднего уха (молоточком и наковальней соответственно). У млекопитающих имеется подъязычная кость с рожками, представляющие собой нижний отдел бывшей подъязычной дуги и первую жаберную дугу. Остальные элементы жаберных дуг становятся хрящами гортани[43].
Позвоночник

Характерные черты в строении позвоночного столба млекопитающих: плоские поверхности позвонков (платицельные позвонки), между которыми располагаются хрящевые диски; чёткое расчленение позвоночника на пять отделов — шейный, грудной, поясничный, крестцовый и хвостовой (только у современных китообразных крестца нет, хотя он имелся у некоторых древних китов[44]); постоянное число шейных позвонков — 7 (лишь у ламантина их 6, а у некоторых ленивцев число шейных позвонков варьирует: у ленивца Гоффмана — от 5 до 6, у трёхпалых ленивцев — от 8 до 10[45])[46]. Первые два шейных позвонка — атлант и эпистрофей — типичного для амниот строения; атлант несёт две сочленовные поверхности для соединения с двумя мыщелками затылочной кости[47]. Грудной отдел позвоночника у млекопитающих состоит чаще всего из 12—15, поясничный — из 2—9, крестцовый — из 1—9 позвонков (чаще всего из четырёх). Наибольшей изменчивостью отличается количество хвостовых позвонков: от 3 у гиббоновых до 49 у длиннохвостого ящера. К передним грудным позвонкам причленяются рёбра, соединённые с грудиной («истинные рёбра» — лат. costae verae) и образующие вместе с ней грудную клетку; остальные грудные позвонки несут не доходящие до грудины «ложные рёбра» (costae spuriae)[48].
Конечности и их пояса
Пояс передних конечностей (плечевой пояс) млекопитающих состоит из двух лопаток и двух ключиц. У некоторых млекопитающих ключиц нет (копытные), у других они плохо развиты либо заменены связками (грызуны, некоторые хищные). У однопроходных имеется ещё воронья кость — коракоид, у большинства же млекопитающих он рудиментарен и прирастает к лопатке в виде клювовидного отростка[49][50].

Передние конечности служат млекопитающим для передвижения по земле, плавания, полёта, хватания. В типичном случае передняя конечность состоит из плеча, которое включает одну плечевую кость, предплечья, образованного локтевой и лучевой костями, и кисти. Локтевая кость служит для сочленения кисти с плечом. Кисть передней конечности состоит из запястья, пясти и пальцев. Запястье состоит из 7 костей, расположенных в два ряда. Число костей пясти соответствует числу пальцев (не более пяти). Пальцы обычно состоят из двух-трёх фаланг (у китообразных число фаланг увеличено, достигает 13—14). Соединяются подвижно сочленённые кости суставами[51][52].
Пояс задних конечностей (тазовый пояс) млекопитающих, как и у других наземных позвоночных, состоит из парных лобковых, седалищных и подвздошных костей (у многих видов они срастаются в одну тазовую кость). Эти три отдела соединяются, образуя вертлужную впадину, к которой причленяется задняя конечность. У яйцекладущих и сумчатых к лобковому отделу примыкают сумчатые кости[53]. У китообразных настоящего таза нет; от тазового пояса у них сохраняются две рудиментарные кости, которые не связаны с осевым скелетом[54].
В состав задней конечности входят бедро, голень (образована большой и малой берцовыми костями) и стопа (в составе которой — предплюсна, плюсна и кости пальцев). У большинства млекопитающих бедренная кость короче голени[51].
Конечности млекопитающих приспособлены к их образу жизни. Так, у наземных форм более длинные бедро и голень; у водных форм, напротив, эти отделы укорочены, а удлинены пясть, плюсна и особенно фаланги пальцев. У рукокрылых развит нормально лишь первый палец передних конечностей, остальные пальцы сильно удлинены; между ними располагается кожистая перепонка. У наиболее совершенных бегунов — копытных — первый палец атрофирован. В случае парнокопытных животное ступает на третий и четвёртый палец, а непарнокопытных — лишь на третий. Скорость передвижения наземных видов сильно варьирует: от 4 км/ч (землеройка) до 105—112 км/ч (гепард)[53].

A — шейный; B — грудной; С — поясничный; D — крестцовый; E — хвостовой.
- 1 — череп;
- 2 — верхняя челюсть;
- 3 — нижняя челюсть;
- 4 — атлант;
- 5 — эпистрофей;
- 6 — лопатка;
- 7 — ость лопатки;
- 8 — плечевая кость;
- 9 — лучевая кость;
- 10 — локтевая кость;
- 11 — фаланги пальцев;
- 12 — кости пясти;
- 13 — кости запястья;
- 14 — грудина;
- 15 — хрящевая часть ребра;
- 16 — рёбра;
- 17 — фаланги пальцев;
- 18 — кости плюсны;
- 19 — кости предплюсны;
- 20 — пяточная кость;
- 21 — малоберцовая кость;
- 22 — большеберцовая кость;
- 23 — надколенник;
- 24 — бедро;
- 25 — седалищная кость;
- 26 — подвздошная кость.
Мускулатура

Высокодифференцированная мышечная система млекопитающих обеспечивает выполнение разнообразных движений и имеет в разных отрядах и семействах существенные особенности в зависимости от характерных способов передвижения. Среди скелетных мышц наибольшее развитие получили мышцы конечностей; перемещение конечностей под тело (у рептилий и ранних синапсид они располагались по бокам) увеличило величину шага и эффективность работы мышц. Характерным анатомическим признаком млекопитающих является также наличие куполообразной мышцы — диафрагмы, которая разделяет грудную и брюшную полости; её возникновение позволило резко интенсифицировать вентиляцию лёгких[55].
У млекопитающих хорошо развита жевательная мускулатура, обеспечивающая захват и механическую обработку пищи, причём у растительноядных форм наиболее сильными являются собственно жевательные мышцы, а у плотоядных — височные мышцы, отвечающие за силу укуса. Для млекопитающих характерна подкожная мускулатура; она участвует в терморегуляции, управляя движением волосяного покрова, обеспечивает свёртывание тела ежей, броненосцев и панголинов в клубок, а также выполняет важную роль при общении животных: приводит в движение вибриссы, отвечает за мимику, особенно развитую у хищных и приматов[56].
Перечисленные виды мышц относят к группе поперечно-полосатых. Поперечно-полосатой мышечной тканью представлены скелетная мускулатура и некоторые мышцы внутренних органов (например, дыхательные мышцы[57] и некоторые сфинктеры[58])[56]. Помимо движения, скелетная мускулатура обеспечивает также сократительный термогенез (подробнее см. ниже). Однако, основная масса стенок внутренних органов образована гладкими мышцами. К гладким относят мышцы желудка, кишечника, стенок кровеносных сосудов, внутренних полых органов (жёлчного пузыря, мочевого пузыря, матки), глаза, выводных протоков желёз внешней секреции и бронхов[59][60]. Особая сердечная мышечная ткань имеется в сердце; в её состав входят сократительные клетки (кардиомиоциты) и проводящие клетки (клетки-пейсмейкеры), которые образуют проводящую систему сердца и генерируют электрические импульсы, управляющие частотой сердечных сокращений[59].
Нервная система
Как и у других позвоночных, нервную систему млекопитающих по морфологическим признакам делят на центральную, включающую головной и спинной мозг, и периферическую, которая слагается отходящими от головного и спинного мозга нервами[61]. В составе центральной нервной системы особо значительное развитие у млекопитающих получил головной мозг: доля его массы в массе всего тела редко бывает ниже 1 %, тогда как у современных рептилий и птиц она составляет от 0,05 до 0,5 %. Увеличение размеров и совершенствование структуры головного мозга позволили ему взять на себя сложные ассоциативные функции, что расширило способности млекопитающих к исследовательскому поведению и накоплению индивидуального опыта[62].
В структурно-функциональном плане нервную систему подразделяют на соматическую, иннервирующую скелетную мускулатуру и органы чувств, и вегетативную, иннервирующую внутренние органы, кровеносные и лимфатические сосуды. Резкого разграничения между данными системами не наблюдают[61].
Особенности нервной ткани
Нервная ткань млекопитающих, как и у других позвоночных, эктодермального происхождения[63] и состоит из нервных клеток (нейронов) и вспомогательных клеток — нейроглии[64]. Нейроны образуют многочисленные контакты (синапсы) друг с другом и осуществляют генерирование и распространение нервных импульсов; таким образом, нейрон — структурная и функциональная единица нервной системы[65].

Передача нервного импульса обусловлена изменениями ионных токов в нейронах, то есть изменениями, затрагивающими ионные каналы нейрона. Для нейронов млекопитающих известно больше 12 типов потенциалозависимых ионных каналов (в то же время в аксонах гигантского кальмара встречается лишь 2 типа ионных каналов)[66]. Глиальные клетки обеспечивают жизнедеятельность нейронов и не участвуют в проведении нервных импульсов[63]. Как и у других позвоночных, в нервной ткани млекопитающих хорошо развито капиллярное кровообращение; саму же нервную ткань подразделяют на серое вещество, образованное телами нейронов и немиелинизированными нервными волокнами, и белое вещество, образованное миелинизированными нервными волокнами[65][64].
Итак, нервный импульс распространяется по нервным волокнам и передаётся от нейрона к нейрону посредством особого рода межклеточных контактов — синапсов. У млекопитающих передача между нейронами иногда может осуществляться с помощью электрических сигналов[67] или, в большинстве случаев, с помощью особых химических соединений-посредников. Эти химические соединения называются нейромедиаторами (в некоторых источниках отдельно рассматривают также нейромодуляторы, отличающиеся от нейромедиаторов тем, что выбрасываются в синаптическую щель в меньших количествах и действуют медленнее[68]). Они связывают специфические рецепторы на постсинаптической мембране, которые могут быть как частью ионного канала, непосредственно влияя на его проницаемость для ионов (ионотропные рецепторы), так и не связанными с ним непосредственно — метаботропные рецепторы; в последнем случае действие нейромедиатора на ионный канал опосредовано различными соединениями — вторичными посредниками, например, цАМФ[69].
Головной мозг

Относительный размер головного мозга млекопитающих очень велик: по массе он в 3—15 раз превосходит спинной мозг (у рептилий их масса примерно одинакова)[70]. Головной мозг млекопитающих включает пять отделов: передний (или конечный), промежуточный, средний, продолговатый мозг и мозжечок, из них особенно развиты первый и последний[71]. В свою очередь, в переднем мозге особенно сильно развита кора больших полушарий, состоящая из серого вещества и содержащая высшие ассоциативные центры.
В то же время, полосатые тела, в которых расположены эти центры у пресмыкающихся и птиц, составляя бо́льшую часть их переднего мозга, у млекопитающих относительно невелики[72]. Полосатые тела выполняют у млекопитающих функцию регуляции инстинктивных реакций, подчиняясь при этом контролю коры полушарий[73][74].
Разросшаяся кора полушарий млекопитающих образовала вторичный мозговой свод (неопаллиум), который накрыл собой весь мозг, а первичный мозговой свод (архипаллиум) был оттеснён внутрь, образовав складку — гиппокамп. Неопаллиум служит у млекопитающих центром высшей нервной деятельности, координирующим работу других отделов мозга. Полушария связаны друг с другом мозолистым телом из белого вещества. Таким образом, по сравнению с птицами, у которых крыша переднего мозга развита относительно слабо и представлена тонким слоем нервных клеток архипаллиума и неопаллиума, крыша переднего мозга развита в значительно большей мере[73][74]. Кора мозга образована несколькими (до 7) слоями тел нервных клеток и покрывает весь передний мозг. У низших представителей класса млекопитающих поверхность коры гладкая, но у высокоорганизованных форм она образует складки и извилины с глубокими бороздами; среди них наиболее чётко обозначены сильвиева борозда, отграничивающая лобную долю коры от височной, роландова борозда, разделяющая лобную и теменную доли, и др.[71] Другие части переднего мозга имеют у млекопитающих относительно меньшие размеры, но сохраняют своё значение. Впереди полушарий лежат обонятельные доли, наиболее развитые у животных с хорошим обонянием.
Эпифиз, гипофиз и гипоталамус, входящие в состав промежуточного мозга, невелики и не видны сверху, однако они ответственны за ряд жизненно важных функций: регуляция метаболизма, поддержание гомеостаза, регуляция эндокринных функций, а также сезонной ритмики функций тела, теплоотдачу. В среднем мозге выделяется четверохолмие, бугры которого являются зрительными и слуховыми центрами; в этом же отделе мозга располагается (как и у других амниот) двигательный центр — красное ядро[75]. Мозжечок отвечает за координацию движений; его поверхность, как и у переднего мозга, имеет много складок и образована серым веществом. В продолговатом мозге локализованы центры дыхания, кровообращения, пищеварения и др.[76]
Среди всех групп позвоночных млекопитающие выделяются наивысшим уровнем развития интеллекта, что связано прежде всего с прогрессивным развитием коры полушарий переднего мозга, хотя этот уровень существенно различается у млекопитающих разных систематических групп. Так, у различных групп млекопитающих отношение массы полушарий переднего мозга к массе всего головного мозга варьирует: у ежей — 48 %, у белок — 53 %, у волков — 70 %, у дельфинов — 75 %[77]. Различным оказывается также и характерный для этих групп уровень развития высшей нервной деятельности. Наивысшего среди млекопитающих развития многие аспекты высшей нервной деятельности достигают у хищных, китообразных, высших приматов[78]. Впрочем, большинство исследователей при оценивании интеллекта различных животных отдают первенство не нейроанатомическому, а функциональному подходу[79].
Установлено, что у большинства млекопитающих сигнальные системы не имеют принципиальных отличий от подобных систем других позвоночных, но существенно более высокий уровень демонстрируют узконосые обезьяны, особенно человекообразные)[80][81].
Спинной мозг

В спинном мозге млекопитающих серое вещество образует в поперечном сечении фигуру «бабочки» с боковыми выступами — «рогами». Хорошо развиты восходящие пути (образованы чувствующими (афферентными) волокнами), по которым информация от частей тела достигает головного мозга. Имеются также нисходящие пути, сформированные двигательными (эфферентными) волокнами. Кроме того, у млекопитающих имеется особый пирамидный путь, идущий от двигательной зоны коры больших полушарий прямо к эфферентным нейронам продолговатого и спинного мозга. При этом волокна латеральной части этого пути перекрещиваются и иннервируют противоположную сторону тела (то есть левое волокно иннервирует правую часть тела, и наоборот), а вентральные волокна сохраняют связь со своей стороной тела[82].
В плечевой и поясничной областях спинной мозг млекопитающих образует утолщения, развитые тем сильнее, чем сильнее развита соответствующая пара конечностей (так, у кенгуру с их мощными задними ногами особо развито поясничное утолщение, а у летучих мышей, передвигающихся при помощи передних конечностей — плечевое). Основная функция спинного мозга — осуществление простых безусловных рефлексов[83].
Периферическая нервная система

Периферическую нервную систему млекопитающих образуют черепные и спинномозговые нервы, отходящие соответственно от головного и спинного мозга, а также их ответвления (всего их насчитывают свыше 200). У млекопитающих, как и у других амниот, имеется 12 пар черепных нервов; число спинномозговых нервов значительно больше, у человека — 31 пара[84].
Большинство (11 пар из 12) черепных и часть спинномозговых нервов функционально относят к соматической нервной системе, они иннервируют кожу, суставы, скелетные мышцы и органы чувств, отвечая за координацию движений тела и восприятие внешних стимулов. Данная система у млекопитающих хорошо развита, обеспечивая, в частности, наивысшую скорость рефлексов[84].
Другая часть спинномозговых нервов, а также в значительной степени блуждающий нерв функционально относят к вегетативной нервной системе. Волокна входящих в неё нервов образованы двумя нейронами. Первый из них находится в центральной нервной системе, его аксон (преганглионарное волокно) передаёт импульс вегетативному ганглию, а уже от него возбуждение по постганглионарному волокну достигает органа-эффектора.
Морфологически и функционально разделяют симпатическую и парасимпатическую вегетативную нервную систему. В случае первой преганглионарные волокна короче постганглионарных, а вегетативные ганглии расположены недалеко от позвоночника; центры локализованы только в спинном мозге. Для парасимпатической системы ситуация обратная: преганглионарные волокна длинны, а постганглионарные волокна коротки, поскольку ганглии лежат недалеко от иннервируемых органов или даже в их стенках; центры расположены в крестцовом отделе спинного мозга и в головном мозге. При этом если у рыб симпатическая и парасимпатическая системы делят органы между собой, то у четвероногих (и, в частности, у млекопитающих) их области иннервации накладываются друг на друга вплоть до полного взаимного перекрывания[85].
Органы чувств
Набор органов чувств у млекопитающих — такой же, как и у других амниот. Органы чувств служат для восприятия поступающих из внешнего мира сигналов различных модальностей; при этом каждой модальности отвечает своя сенсорная система. Органы чувств содержат рецепторы, чья реакция на стимул приводит к генерации нервного импульса, и входят в периферическую часть соответствующей сенсорной системы — наряду с промежуточной (нейронные проводящие пути, по которым передаётся нервный импульс) и центральной (участок коры больших полушарий, ответственный за обработку получаемых сигналов) её частями. Помимо этого, имеются также интероцептивные сенсорные системы, воспринимающие информацию о состоянии внутренней среды организма[86].
Органы зрения

- 1
- склера,
- 2
- сосудистая оболочка,
- 3
- канал Шлемма,
- 4
- корень радужной оболочки,
- 5
- роговица,
- 6
- радужная оболочка,
- 7
- зрачок,
- 8
- передняя камера,
- 9
- задняя камера,
- 10
- цилиарное тело,
- 11
- хрусталик,
- 12
- стекловидное тело,
- 13
- сетчатка,
- 14
- зрительный нерв,
- 15
- цинновы связки.
В состав зрительной сенсорной системы млекопитающих входят органы зрения — глаза (периферическая часть системы), цепи вставочных нейронов с подкорковыми зрительными центрами (промежуточная часть, ответственная за передачу нервных импульсов) и центральная часть — зрительный центр в коре головного мозга[86].
Органы зрения у млекопитающих развиты, как правило, достаточно хорошо, хотя в их жизни они имеют меньшее значение, чем у птиц: обычно млекопитающие обращают мало внимания на неподвижные предметы, так что к стоящему без движения человеку даже столь осторожные звери, как лисица или заяц, могут подойти вплотную. Особенно крупные глаза имеют ночные звери и животные, обитающие в открытых ландшафтах. У лесных зверей зрение не столь острое, а у роющих подземных видов глаза в большей или меньшей мере редуцированы, в некоторых случаях (сумчатые кроты, слепыш, слепой крот) даже затянуты кожистой перепонкой[87][88].
У многих млекопитающих хорошо развито бинокулярное зрение, основанное на формировании двух изображений, полученных каждым глазом, и их последующем сопоставлении. В ходе обмена информацией между обоими зрительными центрами два полученных изображения сливаются в одну трёхмерную картину[89].
Как и у других позвоночных, глаз млекопитающего развивается из переднего мозгового пузыря и имеет округлую форму (глазное яблоко). Снаружи глазное яблоко защищено белковой фиброзной оболочкой, передняя часть которой прозрачна (роговица), а остальная — нет (склера). Следующий слой — сосудистая оболочка, спереди переходящая в радужную оболочку с отверстием в центре — зрачком[90].
Через зрачок отражённый от объектов свет проникает внутрь глаза. Количество пропускаемого света определяется диаметром зрачка, просвет которого автоматически регулируется мышцами радужной оболочки. Хрусталик фокусирует прошедшие через зрачок лучи света на сетчатке — внутреннем слое оболочки глаза, содержащем фоторецепторы — светочувствительные нервные клетки. Окружающие хрусталик мышцы обеспечивают аккомодацию глаза. У млекопитающих для достижения высокой резкости изображения хрусталик при наблюдении близких объектов принимает выпуклую форму, при наблюдении удалённых — почти плоскую[91]. У пресмыкающихся и птиц аккомодация, в отличие от млекопитающих, включает не только изменение формы хрусталика, но и изменение расстояния между хрусталиком и сетчаткой. У мелких грызунов (полёвки, мыши) из-за незначительности обзора способность к аккомодации практически утрачена[92].

Среди фоторецепторов выделяют две основные разновидности — палочки и колбочки, причём палочки преобладают; так, у человека сетчатка содержит около 123 млн палочек и 7 млн колбочек[93]. Палочки отвечают только за восприятие света и обеспечивают ночное зрение, а при дневном зрении ведущую роль играют колбочки, позволяя животным не только воспринимать свет, но и различать цвета[94].
Впрочем, у млекопитающих цветовое зрение развито слабее, чем у птиц с их четырёхкомпонентным зрением: у подавляющего большинства млекопитающих зрение — двухкомпонентное, а трёхкомпонентное цветовое зрение имеется только у высших приматов (узконосые и частично широконосые обезьяны)[95]. Так, европейская рыжая полёвка различает лишь красный и жёлтый цвета, однако кошка способна различать 25 оттенков серого. У опоссума, лесного хоря и некоторых других видов цветное зрение вообще не обнаружено[96]. В то же время некоторые сумчатые, рукокрылые и грызуны способны видеть в ультрафиолетовом диапазоне[97].
Сигнал, воспринятый от сетчатки, поступает в центральную часть зрительной сенсорной системы — зрительную кору (расположена в затылочной доле коры больших полушарий) через зрительный нерв, верхние бугры четверохолмия и латеральное коленчатое тело; данные структуры входят в промежуточную часть зрительной сенсорной системы[98].
Органы слуха и равновесия

Слух и звуковая сигнализация у млекопитающих обслуживают важнейшие жизненные явления — поиск пищи, распознавание опасностей, опознавание особей своего и чужих видов, причём различные отряды отличаются разными особенностями слуха. Млекопитающие, хотя и меньше, чем птицы, обладают звуковой локацией; они издают звуки голосовыми связками и резонируют их воздухоносными путями. Наиболее обычными издаваемыми ими звуками являются рёв, стон, мяуканье, шипенье[99].
Некоторые землеройки и морские львы, зубатые киты и в особенности летучие мыши обладают способностью к эхолокации, издавая и воспринимая ультразвуки; у летучих мышей генератором локационных сигналов служат ротовой аппарат и нос, у зубатых китов — система воздушных мешков надчерепного звукового прохода. Землеройки при эхолокации издают импульсы продолжительностью 5—33 мс с частотой 30—60 кГц. Морской лев способен издавать звуки частотой 20—72 кГц, дельфины — 120—200 кГц, что позволяет им определять положение косяков рыб с расстояния до 3 км[99][100]. У летучих мышей ультразвуковая локация почти полностью заменяет слабо развитое зрение, однако ни один вид летучих мышей не лишён зрения целиком[101]; более того, зрение используется ими для навигации, особенно на дальних расстояниях, когда радиуса действия эхолокации не хватает[102]. Человек в норме может слышать звуки в диапазоне от 20 до 20000 Гц; собаки и лошади могут слышать ультразвуки, а усатые киты и слоны общаются друг с другом инфразвуками[21].
Органы слуха — уши — у подавляющего большинства видов млекопитающих развиты очень хорошо. Ухо делится на три части: внутреннее, среднее и наружное ухо. Если внутреннее и среднее ухо имеются и у других четвероногих, то наружное ухо, включающее ушную раковину, наружный слуховой проход и барабанную перепонку, представляет собой новоприобретение млекопитающих, облегчающее улавливание звуковых волн. Впрочем, у многих водных и роющих видов ушная раковина вторично исчезает[103][104].
Поскольку у позвоночных ухо содержит также вестибулярный аппарат, реализующий чувство равновесия, то более точно именовать слуховую сенсорную систему термином статоакустическая сенсорная система. В её состав, помимо периферической части, которую образуют уши, входят промежуточная часть, ответственная за передачу нервных импульсов и включающая цепи вставочных нейронов вместе с подкорковыми слуховыми центрами, и центральная часть, которая представлена нейронами коркового слухового центра, расположенного в коре головного мозга[105][106].

- Наружное ухо
- 1 — череп;
- 2 — слуховой канал;
- 3 — ушная раковина.
- Среднее ухо
- 4 — барабанная перепонка;
- 5 — овальное окно;
- 6 — молоточек;
- 7 — наковальня;
- 8 — стремечко.
- Внутреннее ухо
- 9 — полукружные каналы;
- 10 — улитка;
- 11 — нервы;
- 12 — евстахиева труба.
Полость среднего уха заполнена воздухом и отграничена от наружного слухового прохода тонкой мембраной — барабанной перепонкой, колеблющейся под воздействием пришедшей через наружный слуховой проход звуковой волны. В данной полости располагаются три слуховые косточки: стремечко, молоточек и наковальня (гомологи соответственно сочленовной и квадратной костей)[70]. Основание стремечка заходит в овальное окно, закрытое мембраной; от него начинается внутреннее ухо. Эти косточки составляют в совокупности рычажную систему, которая передаёт колебания перепонки, усиливая их, на мембрану овального окна внутреннего уха[107].
Во внутреннем ухе, которое расположено в каменистой части височной кости, выделяют вестибулярный и слуховой отделы[108].
Вестибулярный отдел, служащий органом равновесия, включает три полукружных канала, а также два отолитовых органа — мешочек и маточку. Полукружные каналы располагаются в трёх взаимно перпендикулярных плоскостях и заполнены эндолимфой. Они соединяются в полости преддверия, где расширяются в виде ампул. В ампулах имеются рецепторы вестибулярного аппарата — особые волосковые клетки, снабжённые пучками волосков. Каждый пучок сверху покрыт купулой (желеобразным веществом, скреплённым фибриллярными волокнами). При начале или конце вращательного движения эндолимфа начинает давить на купулу, стремясь сместить её в сторону, противоположную направлению движения, и смещение купулы возбуждает волосковые клетки. В полости преддверия также имеются два расширения — мешочек и маточка, сообщающиеся с помощью Y-образного протока. В каждом из расширений имеется возвышение — макула, покрытая отолитовой мембраной, которая содержит множество отолитов, или оокиниев — мелких, но тяжёлых кристаллов карбоната кальция. Макула маточки располагается горизонтально, а макула мешочка — вертикально. Под действием силы тяжести или линейного ускорения отолитовые мембраны сдвигаются относительно макул, раздражая волосковые клетки[109][110].
С волосковыми клетками связаны нейроны вестибулярного ядра, активность которых меняется под влиянием нейромедиаторов, выделяемых волосковыми клетками. Их аксоны формируют один из корешков преддверно-улиткового нерва, который следует к вестибулярным ядрам. От них информация передаётся в таламус и мозжечок, а из них — в кору больших полушарий[110].

В состав слухового отдела внутреннего уха входит улитка — заполненная жидкостью перепончатая трубка, по всей длине которой проходят три параллельных канала: два наружных — вестибулярная лестница) и барабанная лестница — и заключённый между ними более узкий улитковый канал[111][112]. Последний отделён от барабанной лестницы базилярной мембраной, а внутри улиткового канала размещена текториальная мембрана[113]. Вдоль всей длины улитки тянется расположенный на базилярной мембране кортиев орган — эпителиальное образование из двух видов клеток: волосковых клеток, непосредственно отвечающих за восприятие звуковых стимулов, и опорных клеток различного типа, поддерживающих структурную целостность кортиева органа[114][115]. Кортиев орган присущ только млекопитающим; у других четвероногих аналогом кортиева органа служит базилярный сосочек[116][117].
При наличии звукового сигнала вибрации от молоточка, расположенного напротив овального окна, передаются перилимфе — жидкости, заполняющей вестибулярную и барабанную лестницы. Движущаяся волна перилимфы доходит до конца улитки и заворачивает в барабанную лестницу, откуда движется к основанию улитки и доходит до круглого окна. Энергия этой волны заставляет вибрировать базиллярную мембрану, и эта вибрация стимулирует волосковые клетки. Поскольку базиллярная мембрана туго натянута, в каждой её точке наибольший ответ достигается на волны определённой частоты. Каждая волосковая клетка несёт пучок «волосков» (стереоцилий), содержащих внутри актиновые филаменты и погружённых во внутреннюю жидкость средней лестницы — эндолимфу[118][119].
Вибрирование базилярной мембраны вызывает сдвиг текториальной мембраны и последующее отклонение (изгиб) стереоцилий (таким образом, слуховой рецептор представляет собой механорецептор). Из-за этого в мембране волосковой клетки открываются механочувствительные ионные каналы, внутрь клетки входят катионы К+, и клетка деполяризуется. В результате деполяризации открываются кальциевые каналы, внутрь волосковой клетки входит кальций, и она высвобождает нейромедиатор глутамат в синаптическую щель. Это возбуждает афферентный нейрон, и далее сигнал передаётся в ЦНС. Такое возбуждение происходит только при отклонении стереоцилий в одну определённую сторону; это возбуждение сопровождается выделением большего количества глутамата и увеличением частоты генерации потенциала действия афферентным нейроном, однако при отклонении в противоположную сторону имеет место обратный эффект, и клетка гиперполяризуется[118][119].
В центральную часть слуховой сенсорной системы — слуховую кору (расположена в височной доле коры больших полушарий) — сигнал от улитки поступает по следующему пути: преддверно-улитковый нерв, улитковые ядра продолговатого мозга, верхнее оливное ядро варолиева моста, нижние бугры четверохолмия, медиальное коленчатое тело таламуса; данные структуры и составляют в совокупности периферическую часть слуховой сенсорной системы[120].
Органы обоняния
Органы обоняния млекопитающих развиты сильнее, чем у других наземных позвоночных, и играют в их жизни весьма важную роль. О важности обоняния для млекопитающих говорит и тот факт, что наиболее обширное семейство генов в их геноме образуют как раз гены, кодирующие белки обонятельных рецепторов[121]. Способность к различению запахов млекопитающие используют для ориентирования в пространстве, при поисках пищи, в рамках межвидовых и внутривидовых контактов; приятные запахи запускают секрецию слюны и желудочного сока, а неприятные предупреждают о потенциальном вреде (например, неприятный запах испорченной пищи)[122]. По степени развития обонятельной функции млекопитающих подразделяют на две группы: макросматов с исключительно острым обонянием (большинство млекопитающих) и микросматов с умеренно развитым обонянием (приматы, китообразные, ластоногие)[100].

Различие между данными группами хорошо видно при сопоставлении степени развития обоняния у человека и собаки. Если в носу человека имеется около 6 млн обонятельных клеток, то у собаки их — около 225 млн[123]. Многие макросматы чувствуют запахи на расстоянии нескольких сот метров и способны находить пищевые объекты под землёй[124]. Хорошо известна практика поиска растущих в лесу под землёй трюфелей при помощи специально обученных поисковых собак и свиней, которые способны учуять грибы на расстоянии до 20 м[125].
Большинство млекопитающих сохраняют якобсонов орган — обособленный отдел обонятельной капсулы, который представляет собой замкнутую полость, сообщающуюся с полостью рта. Этот орган, имеющийся также у двоякодышащих и большинства четвероногих (важнейшие исключения — птицы и крокодилы), служит главным образом для восприятия феромонов. У представителей ряда групп (китообразные, сирены, большинство рукокрылых и узконосые приматы, включая человека) якобсонов орган рудиментарен или полностью утрачен[126][127][128].

Органы обоняния млекопитающих располагаются в задней верхней части носовой полости, где возникает весьма сложная, особенно у макросматов, система обонятельных носовых раковин — тонких костных лепестков, направленных внутрь полости и покрытых обонятельным эпителием, который содержит многочисленные рецепторы запахов. Данные рецепторы способны улавливать содержащиеся во вдыхаемом воздухе молекулы пахучих веществ. Как и рецепторы вкуса, их относят к группе хеморецепторов. Сигналы о наличии пахнущих веществ передаются через обонятельный нерв в соответствующий центр головного мозга — обонятельную луковицу или первичные центры обоняния коры головного мозга. Из последней обонятельные сигналы передаются в гипоталамус, лимбическую систему, ретикулярную формацию и неокортекс[100][122].
Обонятельные рецепторы (обонятельные клетки) представляют собой биполярные нейроны. Кроме обонятельных клеток, связанных с обонятельным нервом, в слизистой оболочке носа имеются также свободные окончания тройничного нерва; они способны реагировать на некоторые агрессивные запахи, например, кислотные или аммиачные испарения. Рецепторные белки располагаются на мембране ресничек. Как отмечалось выше, они кодируются весьма обширным семейством генов, поэтому возможно, что одна обонятельная клетка экспрессирует лишь один из этих генов. Однако не все эти гены могут экспрессироваться; так, у человека экспрессируется около 40 % из них[129].
Органы вкуса

Органы вкуса млекопитающих представлены вкусовыми почками, или сосочками, которые расположены на слизистых оболочках языка, твёрдого нёба, а также глотки и надгортанника[130] и содержат рецепторы вкуса (хеморецепторы). Традиционно считалось, что система восприятия вкусовых ощущений у млекопитающих — четырёхкомпонентная, причём первичными вкусами являются сладкий, солёный, кислый и горький[131][132]. На рубеже XX—XXI вв. выявлен пятый тип рецепторов вкуса, отвечающий за восприятие «мясного» вкуса (умами)[133]. Вкусовые рецепторы млекопитающих находятся во вкусовых почках, представляющих собой видоизменённые эпителиальные клетки. В 2005 году было установлено, что одна чувствительная клетка экспрессирует только один тип рецепторов, а значит, чувствительная лишь к одному из четырёх вкусов[134].

Рецепторы, реагирующие на сладкий и горький вкус, а также умами, метаботропные и связаны с G-белками. Например, у человека имеется свыше 30 типов рецепторов горького вкуса, но только один — для умами и один — для сладкого. Входящие сенсорные импульсы горького вкуса проводятся G-белком α-гастдуцином. Рецептор умами представляет собой метаботропный глутаматный рецептор (mGluR4), стимуляция которого вызывает уменьшение концентрации цАМФ. Кислый вкус ощущается, когда присутствие ионов H+, характерное для кислой среды, ведёт в более частому закрыванию К+-каналов и тем самым деполяризует чувствительную клетку. Солёный вкус обусловлен наличием катионов Na+, К+ и т. д., поскольку они, входя в чувствительную клетку по специфическим ионным каналам, деполяризуют клетку, однако присутствие анионов также играет свою роль[135][134]. Информация от чувствительных клеток собирается лицевым нервом (передние две трети языка), языкоглоточным нервом (задняя треть языка и твёрдое нёбо) и блуждающим нервом (глотка и надгортанник), откуда она поступает в особый пучок в продолговатом мозге. Далее она поступает в таламус, а далее — в соответствующую зону коры больших полушарий[130].
Ранее считали, что сладкий и солёный вкус воспринимаются преимущественно кончиком языка, кислый — его боками, горький — средней частью спинки языка[132]. Тем не менее, имеющиеся к настоящему моменту молекулярные и функциональные данные показывают, что все вкусовые рецепторы распределены по всей поверхности языка и отличаются лишь плотностью своего распределения[135]. Таким образом, никакой «карты языка» не существует, вопреки ошибочным популярным представлениям[136].
Другие органы чувств

Органы осязания млекопитающих представлены как свободными нервными окончаниями, лежащими в эпителии, так и несвободными нервными окончаниями (заключёнными в капсулы из нейроглии или некапсулированными); среди последних одни лежат в эпителии, другие — в дерме. Свободные нервные окончания выступают в качестве терморецепторов, механорецепторов и ноцицепторов (то есть отвечают за восприятие изменения температуры, механических воздействий и болевые ощущения). К инкапсулированным нервным окончаниям относят рецепторы давления (тельца Пачини) и прикосновения (тельца Мейснера), тельца Руффини, совмещающие функции рецепторов растяжения и тепла, и колбы Краузе — рецепторы, реагирующие на холод[137][138]. Некапсулированные нервные окончания представлены тельцами Меркеля, также относящимися к рецепторам прикосновения (в отличие от птиц, у которых они локализуются в дерме, у млекопитающих эти тельца лежат в глубоких слоях эпидермиса)[139][140].
Важную осязательную функцию у млекопитающих выполняют многочисленные волоски, а также вибриссы: рецепторы волосяных фолликулов оперативно реагируют на малейшее отклонение волоса от исходного положения[138][141].
Иногда у млекопитающих имеются и другие виды экстероцепции. Так, у ряда видов (утконос, ехидна, гвианский дельфин имеются электрорецепторы — рецепторы электрического поля (наиболее развита электрорецепция у утконоса, электрорецепторы на клюве которого воспринимают электрическое поле, порождаемое работающими мышцами ракообразных, лягушек, мелкой рыбы и другой добычи[142])[143][144]. У некоторых млекопитающих обнаружена способность воспринимать магнитное поле (в частности, в придаточных пазухах носа человека — клиновидной пазухе и решётчатом лабиринте — найдены частицы железосодержащего минерала магнетита, что свидетельствует в пользу того, что такая способность есть и у человека[145]); в то же время конкретные клетки, играющие роль магниторецепторов, у млекопитающих пока не выявлены[146].
Дыхательная система

Дыхательная система служит для газообмена организма с окружающей средой, обеспечивая поступление кислорода и выведение углекислого газа. У млекопитающих она представлена органами дыхания (лёгкими) и дыхательными путями: верхними и нижними. В систему верхних дыхательных путей входят полость носа, носоглотка и ротоглотка, а частично — и ротовая полость, поскольку она тоже может участвовать в процессе дыхания. Система нижних дыхательных путей состоит из гортани (иногда её относят к верхним дыхательным путям), трахеи и бронхов[147][148].
Из особенностей воздухопроводящих путей стоит отметить усложнение строение гортани. В её основании лежит перстневидный хрящ, передняя и боковая стенки образованы щитовидным хрящом, который имеется только у млекопитающих. По бокам спинной стороны гортани над перстеневидным хрящом находятся парные черпаловидные хрящи, а к переднему краю щитовидного хряща прилегает надгортанник. Наличие щитовидного хряща и надгортанника — отличительный признак млекопитающих, оно не свойственно другим позвоночным. Между перстеневидным и щитовидным хрящами располагаются мешковидные полости, называемые желудочками гортани. Между щитовидным и черпаловидными хрящами лежат голосовые связки — парные складки слизистой оболочки, вибрация которых позволяет издавать различные звуки[149].
У млекопитающих хорошо развиты трахея и бронхи (внелёгочные и внутрилёгочные). В ходе эмбрионального развития бронхи млекопитающих ветвятся, образуя сложное бронхиальное дерево. В процессе многократного разветвления бронхи переходят в терминальные бронхиолы, а те — в ещё более мелкие дыхательные бронхиолы. Последние лежат уже непосредственно в лёгких и переходят в альвеолярные ходы, заканчивающиеся альвеолярными мешочками. На стенках альвеолярных ходов и альвеолярных мешочков располагаются альвеолы — тонкостенные пузырьки, густо оплетённые капиллярами; через них и происходит газообмен. Число альвеол огромно: у хищных с их активным образом жизни — 300—350 млн, у малоподвижных ленивцев — около 6 млн. Таким образом, благодаря наличию альвеол создаётся огромная поверхность для газообмена[149][150]. Диаметр альвеолы составляет 25—30 мкм, это — предельно малая величина, при которой поверхностное натяжение на влажных стенках альвеол не мешает расправлению лёгких. Чтобы диаметр альвеолы не уменьшался ниже допустимого, на них имеется особая липидная плёнка, снижающая поверхностное натяжение — сурфактант[151].
Благодаря наличию альвеол, у млекопитающих лёгкие ячеистые и не продуваются транзитным потоком воздуха, а набирают воздух. Такой принцип дыхания у млекопитающих доведён до совершенства, именно его высокая эффективность позволяет млекопитающим поддерживать высокую скорость обмена веществ, а за счёт её — постоянную температуру тела. Лёгкие не имеют полостей, и избежать возникновения значительных внутренних напряжений в ходе изменения объёма лёгких удаётся благодаря разделению лёгких на доли и возможности взаимного скольжения этих долей[152].
Попеременное изменение объёма лёгких обеспечивают не только движения грудной клетки, но и движения диафрагмы, наличие которой — характерный анатомический признак млекопитающих[149]. Дыханием управляет расположенный в продолговатом мозге дыхательный центр. Повышение в крови (а значит, и в спинномозговой жидкости) концентрации углекислого газа, сопровождающееся образованием угольной кислоты H2CO3, влечёт возрастание концентрации ионов H+; они раздражают нервные клетки, которые и дают сигнал межрёберным мышцам усилить дыхательные движения[153].
Число дыхательных движений, выполняемых млекопитающим за минуту, существенно зависит от его размеров: чем больше отношение поверхности тела к его массе, тем чаще дыхание и интенсивнее обмен[154]. Так, лошадь совершает 8—16 таких движений, чёрный медведь — 15—25, человек — 15—20, лисица — 25—40, серая крыса — 100—150, домовая мышь — около 200[155][156]. Кроме того, частота дыхания зависит от физического состояния животного (покой, движение)[154].
Известно, что многие водные млекопитающие, например, морские котики, морские львы, тюлени, китообразные и другие способны находиться долго под водой. Так, тюлень Уэдделла способен находиться на глубине 200—500 м до 20 минут. Существует ряд приспособлений, позволяющих водным млекопитающим долго находиться под водой, не совершая дыхательных движений. Во-первых, эти млекопитающие очень эффективно запасают кислород. Так, тюлень Уэдделла (Leptonychotes weddellii) запасает примерно в два раза больше кислорода на килограмм массы, чем человек, причём лишь 5 % запасённого кислорода находится в сравнительно маленьких лёгких этого тюленя, а 70 % кислорода находятся в крови (у человека 36 % кислорода находится в лёгких и 51 % — в крови). Оставшиеся 25 % кислорода у тюленя Уэдделла (соответственно, 13 % у человека) находятся в связанной с белком миоглобином форме и локализованы в мышцах. Во-вторых, водные млекопитающие сводят к минимуму число производимых движений и изменяют глубину своего погружения за счёт изменения плавучести, а не мышечной работы. При погружении в воду у них снижается пульс и потребление кислорода, и специальные регуляторные механизмы направляют большую часть крови к головному и спинному мозгу, глазам, надпочечникам и плаценте (в случае беременных животных). Приток крови к мышцам сильно ограничен или вовсе отсутствует. Наконец, когда в мускулах заканчивается весь кислород, связанный миоглобином, то мышцы переходят на анаэробное дыхание[157].
Роль кожи млекопитающих в газообмене незначительна: через кожные кровеносные сосуды поступает лишь около 1 % кислорода. Это объясняется как ороговением эпидермиса, так и тем, что общая поверхность кожи ничтожно мала по сравнению с суммарной дыхательной поверхностью лёгких[154].
Кровеносная система
У млекопитающих хорошо развита кровеносная система, важнейшие функции которой — обеспечение снабжения тканей кислородом и питательными веществами, освобождение их от продуктов распада, транспорт гормонов и иммунных факторов. Центральным органом кровообращения служит сердце, ритмичные сокращения мускулатуры которого заставляют кровь циркулировать по кровеносным сосудам. У млекопитающих сердце четырёхкамерное, что отличает их от большинства рептилий с трёхкамерным сердцем, но роднит с птицами и крокодилами. От сердца млекопитающих отходит единственная дуга аорты — левая (этим они отличаются от крокодилов с двумя дугами аорты и от птиц — с единственной, но правой). Ещё одна особенность млекопитающих — отсутствие ядер у эритроцитов, что повышает их кислородную ёмкость[158][159].
Сердце

Сердце млекопитающих состоит из правого и левого желудочков, а также правого и левого предсердий. Предсердия собирают поступающую к сердцу по венам кровь и направляют её в желудочки, а те, сокращаясь, выбрасывают кровь в лёгочные артерии. Камеры сердца сообщаются между собой и с магистральными сосудами — аортой, лёгочным стволом — при помощи отверстий, снабжённых клапанами. Сердце заключено в околосердечную сумку — перикард[160][161].

Как упоминалось выше, мышечная ткань сердца (миокард) имеет особое строение и представляет собой функциональный синцитий. В ней, помимо клеток, способных к сокращению, — кардиомиоцитов, имеются клетки, неспособные сокращаться, но способные к генерации электрических импульсов и проведению их к сократительным клеткам. Они образуют т. н. проводящую систему сердца[59]. Между её элементами и кардиомиоциами имеются щелевидные контакты (нексусы), а между кардиомиоцитами — особые вставочные пластинки. Низкое электрическое сопротивление нексусов и вставочных пластинок обеспечивает распространение возбуждения с проводящей системы на кардиомиоциты и далее по всему миокарду. Способность клеток проводящей системы (водитедей ритма, или пейсмейкеров) к самостоятельной генерации возбуждения обусловливает автоматизм миокарда. Важнейшими элементами проводящей системы являются синоатриальный узел, расположенный между местом впадения правой полой вены и ушком правого предсердия, атриовентрикулярный узел, локализованный в нижней части межпредсердной перегородки, от которого отходит пучок Гиса, по мере приближения к верхушке сердца распадающийся на волокна Пуркинье[162].
В зависимости от образа жизни и коррелирующего с ним уровня интенсивности обмена веществ у разных видов млекопитающих относительная масса сердца существенно различна. Так, сердечный индекс (измеряемое в процентах отношение массы сердца к общей массе тела) у кашалота и ленивца равен 0,3, у африканского слона — 0,4, у серой полёвки — 0,6, а у ушана и обыкновенной землеройки — 1,4[163].
Средняя частота сокращений сердца в минуту также увеличивается с уменьшением размеров млекопитающего и повышением его двигательной активности. Так, у масковой бурозубки она равна 780, малого бурундука — 680, домовой мыши — 600, обыкновенного ежа — 250, собаки — 140, овцы и человека — 70—80, лошади — 35—55, азиатского слона — 25—50. Эта частота у водных форм значительно снижается после погружения под воду (у тюленя — со 180 до 30—60), что позволяет экономнее расходовать запасы кислорода в лёгких и воздухоносных полостях[164][165].
Кровеносные сосуды

Правая сторона сердца обеспечивает малый круг кровообращения (лёгочный): из правого желудочка венозная кровь поступает по лёгочным артериям в лёгкие, откуда по лёгочным венам обогащённая кислородом кровь следует в левое предсердие. Левая сторона сердца обеспечивает работу большого круга кровообращения: от левого желудочка отходит левая дуга аорты, через ответвления которой артериальная кровь разносится по всему телу. Затем кровь, отдав тканям организма кислород и питательные вещества и забрав углекислоту и продукты обмена, через венозную систему возвращается к сердцу — к правому предсердию, в которое впадают передняя (собирающая кровь от головы и передних конечностей) и задняя (от задней части тела) полые вены[166][167].
Порядок ответвления главных артерий от аорты различен. У большинства видов от аорты первой отходит короткая безымянная артерия, делящаяся на правую подключичную и сонные (левую и правую) артерии, затем — левая подключичная артерия. В других случаях левая сонная артерия самостоятельно отходит от аорты (подключичные артерии снабжают кровью передние конечности, а сонные — голову)[163]. Продолжением аорты служит спинная аорта, от которой ответвляются внутренностная (идёт к органам брюшной полости), почечные (к почкам), а её конечными ветвями является хвостовая артерия (к хвосту) и две подвздошные артерии (к задним конечностям). В конечном счёте все артерии, отходящие от аорты, разветвляются вплоть до множества капилляров, общая площадь сечения которых почти в тысячу раз превышает диаметр аорты[168][169].
В свою очередь, капилляры собираются в вены. Среди крупных вен отметим: яремную (идёт от головы) и подключичные (от передних конечностей) вены; сливаясь, яремная и левая подключичная вены образуют безымянную вену, которая вместе с правой подключичной и правой и левой непарными венами (идут от грудной полости) впадает в переднюю полую вену (у большинства млекопитающих, впрочем, левая непарная вена утрачивает прямую связь с полой веной и — через поперечную вену — впадает в правую непарную вену); подвздошные (идут от задних конечностей), хвостовая (от хвоста), почечные (от почек) и печёночные (от печени) вены, впадающие в заднюю полую вену. Особняком стоит воротная вена, несущая кровь от непарных органов брюшной полости (кишечника, желудка, селезёнки, поджелудочной железы) к печени, в капиллярах которой кровь освобождается от возможных токсинов. Отличительная черта млекопитающих — отсутствие аналогичного воротного кровообращения в почках, имеющегося у других четвероногих[167][163].
Кровь и лимфа




Кровь и связанная с ней лимфа — жидкие соединительные ткани организма млекопитающего[170]. Кровь состоит из плазмы и форменных элементов: тромбоцитов, лейкоцитов и эритроцитов, которые образуются в костном мозге. Тромбоциты (у млекопитающих они, в отличие от других позвоночных, безъядерные кровяные пластинки[171][172]) играют определяющую роль в процессе свёртывания крови, а также участвуют в метаболизме серотонина. Лейкоциты (белые кровяные тельца) — часть иммунной системы организма. Эритроциты (красные кровяные тельца) содержат гемоглобин, обеспечивающий перенос кислорода от лёгких к тканям организма[173].
Высокому уровню метаболизма млекопитающих отвечает большое количество эритроцитов в 1 мм3 крови — 8,4 млн (у рептилий — 1,0 млн, у хвостатых земноводных — 0,1 млн)[156]. Эритроциты млекопитающих отличаются малым размером, уплощённо-дисковидной формой (у мозоленогих — овальной) и отсутствием ядер. Изогнутая форма эритроцитов увеличивает площадь поверхности для диффузии кислорода внутрь эритроцита, а благодаря отсутствию ядра в эритроците помещается больше гемоглобина, связывающего кислород. У эритроцитов также отсутствуют митохондрии, и они синтезируют АТФ исключительно за счёт анаэробного дыхания; если бы эритроциты потребляли кислород в ходе аэробного дыхания, они бы не были такими эффективными его переносчиками[174][175].
Соотношение объёмов форменных элементов крови и плазмы у разных видов млекопитающих варьирует. Так, среднее значение гематокрита (выраженное в процентах отношение объёма, занимаемого форменными элементами крови, к её общему объёму) заметно выше у одних видов (человек — 42, собака — 45, кошка — 40, крыса — 43, мышь — 40, коала — 43, ехидна — 48 %) и ниже у других (корова — 30, овца и свинья — 33 %). При этом с увеличением гематокрита, как правило, повышается вязкость крови, хотя на конкретную величину последней сильно влияют и другие факторы[176][177]. Существенно повышенные значения гематокрита и вязкости крови характерны для морских млекопитающих: у калана гематокрит равен 52,5, северного морского слона — 57, гренландского кита — 59 и тюленя Уэдделла — 63,5 %[178][179].
Лимфатическая система выступает как дополнение к кровеносной системе и служит посредницей в обмене веществ между кровью и тканями. Лимфа по происхождению — межтканевая жидкость, образованная просочившейся через стенки капилляров плазмой крови и протекающая по лимфатическим сосудам. В лимфатической системе млекопитающих отсутствуют «лимфатические сердца» — пульсирующие участки сосудов, имеющиеся у земноводных и пресмыкающихся, поскольку при характерном для млекопитающих подвижном образе жизни движение лимфы по сосудам обеспечивается сокращениями скелетных мышц[168][169]. Форменными элементами лимфы являются лимфоциты, которые могут переходить из крови в лимфу и обратно[169][180].
Иммунная система
Иммунная система позволяет организму противостоять патогенам, а также избавляет его от собственных ненормальных клеток (например, опухолевых[181]). Каждый организм имеет неспецифический (врождённый) иммунитет, который дополняется специфическим (приобретённым) иммунитетом[182]. И неспецифический, и специфический иммунитет имеет несколько уровней защиты, однако неспецифические иммунные реакции протекают быстрее специфических[183].
Несмотря на общее сходство, даже между такими близкими видами, как корова и овца, мышь и крыса, человек и другие приматы, наблюдают различия на уровне иммунных тканей, клеток и растворимых медиаторов. Особенно заметны различия на уровне семейств генов, чьи представители характеризуются большой сложностью и полиморфизмом — например, генов, кодирующих антитела, Т-клеточные рецепторы, главный комплекс гистосовместимости, а также рецепторы натуральных киллеров[184]. Среди млекопитающих иммунная система наиболее хорошо изучена у человека и мыши.
Неспецифический иммунитет
Клетки неспецифического иммунитета





Неспецифический иммунитет является эволюционно более древним[185] и включает несколько барьеров (уровней защиты) против чужеродных агентов.
- Физические барьеры. К физическим барьерам, препятствующим проникновению болезнетворных агентов во внутреннюю среду организма, относят:
- кожу — механический барьер для возбудителей. Кроме того, кожа имеет сниженный pH из-за потовых выделений молочной и жирных кислот, препятствующий развитию микроорганизмов.
- слизистые оболочки дыхательных путей и желудочно-кишечного тракта.
- слизь, секреты слёзных и слюнных желёз[186].
- Физиологические барьеры включают:
- температуру тела. Например, возбудитель сибирской язвы Baccilla anthracis прекращает размножение при температуре 41—42 °C, поэтому организмы с большей температурой тела будут резистентны к сибирской язве.
- pH. Одним из таких факторов является кислая среда желудочного сока.
- растворимые факторы. В естественной невосприимчивости принимают участие такие факторы, как лизоцим — гидролитический фермент, содержащийся в слюне, способен разрушать пептидогликановый слой клеточной стенки бактерий; интерфероны — белки, продуцируемые вирусинфицированными или активированными клетками, одной из их функций является прямое подавление размножения вирусов; система комплемента — группа сывороточных белков, которые циркулируют в неактивной форме, однако способны активироваться различными иммунологическими механизмами специфического и неспецифического характера и в активированном состоянии принимать участие в контролируемом ферментативном каскаде, в результате которого повреждается мембрана бактериальных клеток или наступает их опсонизация[187].
- Клеточные барьеры включают:
- натуральные киллеры — большие гранулярные лимфоциты, лишённые признаков Т- или В-лимфоцитов[188], которые специализируется на неспецифической защите против вирусов, микобактерий, опухолевых клеток и т. д. Они закрепляются к опсонизированным поверхностным антигенам и с помощью специальных агентов (перфоринов) формируют поры в стенках клеток-мишеней, тем самым способствуя их дальнейшему лизису[189]. Таким образом, натуральные киллеры находятся на стыке неспецифического и специфического иммунитета[190].
- фагоциты: нейтрофилы, моноциты, макрофаги и эозинофилы. Макрофаги происходят от моноцитов, мигрирующих в ткани; некоторые макрофаги сохраняют мобильность (свободные макрофаги), тогда как другие ограничены некоторой областью (фиксированные макрофаги). Фиксированные макрофаги находятся, например, в клетках Купфера печени, клетках Лангерганса кожи, синусе селезёнки, лимфоузлах и т. д. Моноциты, циркулирующие в крови, и макрофаги объединяются понятием система мононуклеарных фагоцитов. Фагоциты способны мигрировать и проникать сквозь эндотелий, после чего они повреждают локализованные в тканях микроорганизмы с помощью лизоцима, оксидантов (например, H2O2), кислородных радикалов и оксида азота (NO).
При неспецифических иммунных реакциях, в отличие от специфических, не происходит увеличения численности специальных клеток, ориентированных на защиту от конкретного антигена, поэтому неспецифические защитные механизмы также называют неклональными[185].
Специфический иммунитет
Основу специфического иммунитета составляют Т-лимфоциты (Т-клетки) и В-лимфоциты (В-клетки)[182]. Специфический иммунитет можно подразделить на клеточный и гуморальный.
Клеточно-опосредованный специфический иммунный ответ осуществляется Т-клетками. Он требует участия специализированных антигенпрезентирующих клеток, которыми могут выступать дендритные клетки, макрофаги и В-клетки. В антигенпрезентирующей клетке фрагмент антигена связывается с молекулой главного комплекса гистосовместимости, и образовавшийся комплекс выставляется на поверхность клетки, становясь доступным для распознавания Т-клетками[191]. После распознавания антигена Т-клетка начинает выделять интерлейкин 2, который стимулирует клональную экспансию этих моноспецифических Т-клеток. В ходе клональной экспансии Т-клетки дифференцируются на Т-киллеры (цитотоксические Т-клетки), Т-хелперы I типа и Т-хелперы II типа. Т-киллеры убивают инфицированные клетки при помощи токсичных белков[192], Т-хелперы I типа активируют макрофаги и запускают воспалительный процесс, а Т-хелперы II типа необходимы для активации В-клеток[193].
Гуморальная специфическая иммунная реакция осуществляется В-клетками. Активация их начинается с того, что с многочисленными мономерами иммуноглобулинов IgM и IgD, прикреплёнными к поверхности В-клеток, связывается соответствующий антиген. Однако одного образования поверхностного комплекса антиген-антитело недостаточно; для активации необходим второй сигнал, который может поступать либо от тимуснезависимого антигена (например, бактериального полисахарида), либо тимусзависимого (от Т-хелперов II типа). В последнем случае В-клетка передаёт антиген Т-хелперу. Если Т-хелпер распознаёт этот антиген, то он начинает выделять интерлейкин 4, который стимулирует клональную селекцию специфических В-клеток, которые, дифференцируясь в плазмоциты, производят антитела (иммуноглобулины, Ig) против этого антигена[194]. Механизм действия образуемых антител на чужеродный агент может быть различным. Они могут связывать поверхностные антигены вируса и блокировать его способность к инфицированию других клеток (нейтрализация вирусной частицы), могут связываться с поверхностными антигенами на поверхности бактерии и привлекать к ней макрофагов, запуская фагоцитоз (уже упоминавшаяся опсонизация). Антитела могут также связывать антигены на поверхности бактериальной клетки, а с антителами связывается система комплемента, белок которого образует поры в мембране бактериальной клетке, тем самым убивая её[195]. Выделяют 5 классов антител: IgG — основной класс антител крови; IgM — первые антитела, продуцируемые при развитии иммунного ответа; IgA — основной класс антител, содержащихся в молоке, слизи, слезах, секретах дыхательных путей и кишечника; IgD — функционируют как рецепторы клеточных мембран; IgE — осуществляют защиту от паразитов, обусловливают развитие аллергических реакций (подробнее об аллергии см. ниже)[196].
При столкновении с антигеном образуются также Т- и В-клетки памяти, которые могут при повторном столкновении с этим антигеном быстро дифференцироваться соответственно в Т-киллеры или плазмоциты и начать производить антитела[197].
Описанные выше механизмы представлены на иллюстрациях:
-
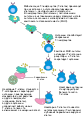
Механизм активации Т-лимфоцитов
-
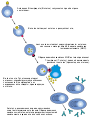
Механизм активации В-лимфоцитов
-
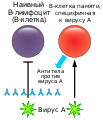
Механизм работы В-клеток памяти



Органы иммунной системы
Все клетки крови образуются из плюрипотентных стволовых клеток костного мозга. Однако в определённый момент эмбрионального развития часть из них мигрирует в тимус (вилочковую железу) или остаются в костном мозге и начинают созревание в Т- и В-лимфоциты, соответственно (по этой функции костный мозг млекопитающих аналогичен фабрициевой сумке птиц). В связи с этим тимус и костный мозг называют первичными лимфоидными органами. В них продолжается клеточная пролиферация и дифференциация. В процессе созревания Т-лимфоциты контактируют со специализированными эпителиальными клетками, дендритными клетками и макрофагами, что даёт возможности для отбора Т-клеток, нужных иммунной системе в этот момент. Важную роль в созревании Т-клеток играют цитокины и интерлейкины 1, 2, 6 и 7. В ходе созревания многие лимфоциты, особенно направленные на распознавание белков самого организма, подвергаются разрушению. Созревание В-клеток также зависит от цитокинов и интерлейкинов 1, 6, 7[198].
Зрелые Т- и В-лимфоциты покидают места своего созревания и мигрируют во вторичные лимфоидные органы: селезёнку, лимфатические узлы, а также лимфоидную ткань слизистых оболочек (англ. mucosa-associated lymphoid tissue, MALT). Она представляет собой скопления лимфоидных клеток, расположенные под слизистой желудочно-кишечного тракта, дыхательных путей, мочевыводящих путей, слёзных желёз. Кроме того, по всему телу разбросаны беспорядочно разбросаны лимфатические клетки, иногда организованные в крупные скопления, например, миндалины или пейеровы бляшки[198].
Аллергия
Аллергия (реакция гиперчувствительности) представляет собой специфический преувеличенный иммунный ответ на чужеродное (обычно безвредное) вещество или антиген (антигены, вызывающие аллергию, называют аллергенами). При первом контакте с антигеном происходит сенсибилизация (аллергизация), а при последующих контактах происходит разрушение здоровых клеток и интактных тканей из-за воспалительных реакций. Аллергия широко распространена среди млекопитающих, в частности, у человека и распространённых домашних животных: кошек, собак, лошадей[199]. Наиболее распространены аллергические реакции, образуемыми в ответ на антиген антителами IgE, которые активно связываются с тучными клетками. В результате тучные клетки быстро высвобождают наиболее активные медиаторы воспаления (гистамин, лейкотриены и др.), в результате чего в течение нескольких минут развивается иммунная реакция (анафилаксия). Из-за быстроты развития реакции этот тип аллергических реакций получил название гиперчувствительности немедленного типа[200].
Иммунологическая толерантность при беременности
При беременности у самок эутериев наблюдается иммунологическая толерантность по отношению к плаценте и развивающемуся плоду[201]. Это явление можно рассматривать как пример необычно успешной аллотрансплантации, поскольку плод и плацента генетически отличаются от организма матери[202]. При этом следует иметь в виду, что при беременности подавляются два иммунных ответа: местный иммунный ответ на плаценту со стороны матки и системный ответ на клетки зародыша со стороны всего материнского организма[203]. Существует множество механизмов, обусловливающих этот «иммунологический парадокс» беременности. Некоторые из них являются плацентарными механизмами, то есть обеспечивают функционирование плаценты как иммунологический барьер между матерью и плодом, создавая иммунологически привилегированную зону. Существует также гипотеза фетоэмбрионической защитной системы эутериев (англ. Eutherian fetoembryonic defense system (eu-FEDS)), согласно которой растворённые в цитоплазме и заякоренные в мембране гликопротеины, экспрессирующиеся в гаметах, подавляют любой иммунный ответ на плод или плаценту[204]. Тем не менее, многие случаи спонтанных абортов обусловлены именно иммунным ответом организма матери на плод[202]. Другим примером, демонстрирующим недостаточность иммунологической толерантности при беременности, является резус-конфликт; кроме того, одной из причин преэклампсии является ненормальный иммунный ответ матери на плаценту[205].
Терморегуляция

Млекопитающих относят к гомойотермным (теплокровным) животным, обеспечивающим определённый уровень температуры тела в основном за счёт внутренних физиолого-биохимических процессов (исключение составляет пойкилотермный грызун голый землекоп[206]). При этом у них хорошо развита терморегуляция: интенсивность процессов, обеспечивающих выделение тепла, регулируется рефлекторно. Традиционно принято считать, что температура тела у млекопитающих находится под контролем сигналов управляющего центра терморегуляции, расположенного в гипоталамусе[207]. Впрочем, согласно современным представлениям, концепция единого центра терморегуляции отвергается и считается, что температура тела регулируется различными термоэффекторными путями, каждый из которых имеет собственные афферентные и эфферентные ветви[208]. Назначение системы терморегуляции — поддержание постоянного значения температуры тела. В ходе эволюции млекопитающие выработали разнообразные средства терморегуляции, которые могут осуществляться на уровне нервной и гуморальной регуляции и затрагивать метаболизм, энергетический обмен и поведение животного. При этом активация тех или иных механизмов зависит от времени суток, сезона, пола и возраста животного[209]. Ниже перечислены некоторые из этих стратегий и механизмов.
От излишней потери тепла млекопитающих предохраняет шёрстный покров, а водные формы — подкожный слой жира. Излишки тепла отводятся потоотделением. Значительную роль в терморегуляции играет развитая система кожных кровеносных сосудов, диаметр просветов которых регулируется нервно-рефлекторным путём[210]. Наличие в носовой полости млекопитающих носовых раковин вынуждает воздух при дыхании проходить по узким носовым ходам между этими раковинами, уменьшая потери тепла и влаги[207]. Среди современных позвоночных такие раковины, помимо млекопитающих, имеют только птицы, так что палеонтологи считают их наличие остеологическим коррелятом гомойотермии. Важнейшим доводом в пользу того, что гомойотермными были уже ближайшие предки млекопитающих, стало обнаружение выступов на внутренней поверхности носовых и верхнечелюстных костей тероцефалов и цинодонтов — свидетельство наличия носовых раковин и у этих продвинутых терапсид[41].
Выделяют два способа теплообразования: сократительный термогенез, при котором теплообразование обусловлено сокращениями скелетных мышц (частный случай — холодовая мышечная дрожь), и несократительный термогенез, когда активизируются процессы клеточного метаболизма: липолиз (в частности, бурой жировой ткани), а также гликогенолиз и гликолиз[211]. Уровень метаболизма у млекопитающих в несколько раз выше, чем у пресмыкающихся, но не столь высок, как у птиц, у которых выше и температура тела[212].
У большинства млекопитающих температура тела относительно постоянна, диапазон её суточных колебаний составляет примерно 1—2 °C. Средние значения температуры тела составляют 30 °C у однопроходных, 36 °C у сумчатых и 38 °C у плацентарных[213].
Отличие температуры тела от температуры внешней среды может достигать 100 °C: у песца даже при 60-градусном морозе температура тела сохраняет постоянное значение — примерно +39 °C. Постоянство температуры, однако, не характерно для однопроходных и сумчатых, а также мелких плацентарных: у однопроходных она меняется в пределах от 22 до 37 °C, у сумчатой крысы — от 29,3 до 37,8 °C, у тенреков — от 13 до 34 °C, у обыкновенной полёвки — от 32 до 37 °C[214]; у двупалых ленивцев она колеблется от 24 до 33 °C[215]. Исключительно велик и уникален для теплокровных диапазон колебаний температуры тела у гладконосых летучих мышей: они способны сохранять жизнеспособность при изменении температуры тела в пределах от −7,5 до +48,5 °C[216].
Химическая терморегуляция у ряда видов млекопитающих устанавливается уже в первые дни после рождения. У других (в частности, у мелких землероек и грызунов[36]) — значительно позже, так что их детёныши на какое-то время оказываются пойкилотермными; они согреваются родителями или же укрываются от холода в гнёздах или норах[217].
Пищеварительная система
В состав пищеварительной системы млекопитающих входят пищеварительный тракт, включающий 4 отдела — рото-глоточную полость, пищевод, желудок, кишечник — и пищеварительные железы[218].
Пищеварительная система млекопитающих отличается значительной сложностью. Она характеризуется общим удлинением пищеварительного тракта, его большей, по сравнению с другими группами позвоночных, дифференцировкой и большей развитостью пищеварительных желёз[219].
Полость ротоглотки


Ротовое отверстие у млекопитающих ограничено присущими только им мягкими подвижными губами; такие губы, впрочем, у однопроходных отсутствуют, а у китообразных слабо развиты и лишены подвижности. Между губами и зубами располагается преддверие рта — полость, которая у утконоса, некоторых сумчатых, многих грызунов и обезьян включает защёчные мешки, служащие для временного хранения и переноса корма[220][221].
После попадания в ротовую полость пища пережёвывается зубами, которые имеются у большинства млекопитающих, кроме современных однопроходных, усатых китов, панголинов и муравьедов. Они находятся в ячейках (альвеолах) челюстных костей. Различают корень зуба, его шейку и выступающую наружу коронку. Зубы млекопитающих дифференцированы на резцы, клыки, предкоренные и истинные коренные. Резцами млекопитающие откусывают пищу, клыками схватывают её и удерживают (иногда — убивают добычу), предмолярами и молярами размельчают. Обычна смена зубов с возрастом, когда т. н. молочные зубы замещаются постоянными (у сумчатых такой смены зубов нет). У грызунов, зайцеобразных, вомбатов резцы (а у части видов — и коренные зубы) растут в течение всей жизни, стачиваясь неравномерно и за счёт этого оставаясь острыми[222][223].
Количество зубов, их форма и размер существенно отличаются у разных млекопитающих; во многом это объясняется характером их питания. У достаточно примитивных насекомоядных зубы слабо дифференцированы. Для грызунов характерны сильно развитая единственная пара резцов, служащая для срезания или обгрызания растительности, отсутствие клыков и плоская жевательная поверхность коренных зубов, предназначенная для перетирания пищи. У хищных сильно развиты клыки, приспособленные для захвата и умерщвления добычи; коренные зубы имеют режущие вершины и плоские жевательные выступы[224].
Характеристику зубной системы млекопитающих можно компактно записать в виде зубной формулы, в которой указывается количество различных зубов в половине верхней и нижней челюсти (в числителе и знаменателе дроби соответственно). Например, зубная формула волка такова:
- [math]\displaystyle{ I{3 \over 3} C{1 \over 1} P{4 \over 4} M{2 \over 3}\,\,, }[/math]
где [math]\displaystyle{ I }[/math] — резцы (incisivi), [math]\displaystyle{ C }[/math] — клыки (canini), [math]\displaystyle{ P }[/math] — предкоренные (praemolares) и [math]\displaystyle{ M }[/math] — истинные коренные (molares) зубы[225].
Если у верхнетриасового морганукодона имелось 54 зуба[226], то у современных видов их число меньше. Максимальное число зубов среди современных сумчатых имеет намбат (Myrmecobius fasciatus): 50—52, среди ныне живущих плацентарных — большеухая лисица (Otocyon megalotis): 48[227]; у большинства мышиных — 16 зубов, у собаки — 42, кошки — 30, узконосых приматов — 32[228]. Эти подсчёты не касались зубатых китов и броненосцев, зубы которых утратили гетеродонтность. У зубатых китов зубы имеют простую коническую форму[229]; число таких зубов обычно весьма велико, а наибольшее значение составляет у ла-платского дельфина (Pontoporia blainvillei): от 210 до 240! У броненосцев зубы одинаковой цилиндрической формы, лишённые эмали и корней; их количество (от 8 до 100) оказывается различным не только у разных видов, но и у разных особей одного и того же вида[230].
У многих млекопитающих имеется диастема — больший, по сравнению с обычным, промежуток между зубами; он, как правило, возникает при редукции части зубов. Диастемы такого происхождения имеются в верхней и нижней челюсти у грызунов, лошадей, кенгуру и других. У жвачных диастема сохраняется лишь в нижней челюсти. Для хищных характерна диастема, возникающая без редукции зубов[231].
При пережёвывании пищи она смачивается слюной, поступающей по протокам из слюнных желёз, что облегчает её проглатывание и продвижение по пищеводу. Ферментативная обработка пищи осуществляется гидролитическими ферментами, содержащимися в слюне — прежде всего α-амилазой, под действием которой сложные углеводы (крахмал, сахар), содержащиеся в пище, распадаются на мономеры, а также липазой, рибонуклеазой и протеазами[232]. Слюнные железы сильно развиты у травоядных животных. Корова, например, выделяет в сутки около 56 л слюны[233]. Слюна млекопитающих содержит не только пищеварительные ферменты, но и бактерицидный белок — лизоцим, обладающий выраженными антисептическими свойствами[234]. Кроме того, слюна является основным источником кальция, поступающим в эмаль зубов, а её щелочная среда защищает ротовую полость от кислых продуктов жизнедеятельности микроорганизмов[232]. У некоторых насекомоядных (короткохвостая бурозубка, водяная кутора, щелезубы) слюна содержит вырабатываемый подчелюстной железой токсин, способный обездвиживать их жертв — мелких грызунов[235][236]. Наконец, слюна облегчает процессы жевания и глотания пищи и составляет 10—20 % от количества пищи, поступающей в пищеварительный тракт[232].
В перемещении и проглатывании пищи участвует язык, служащий также органом вкуса и осязания, а у человека — и ведущим артикуляционным органом при произнесении звуков членораздельной речи. Из полости рта пища попадает в глотку; при глотании мягкое нёбо рефлекторно перекрывает проход в носовую полость, а вход в гортань блокируется движением надгортанника[234].

Пищевод
Пищевод представляет собой тонкостенную трубку, по которой комок пищи попадает в желудок. У большинства млекопитающих пищевод снабжён гладкой мускулатурой, сокращения которой обеспечивают продвижение пищи к желудку. Лишь у жвачных пищевод имеет поперечно-полосатые мышцы, с помощью которых пища («жвачка») отрыгивается из желудка обратно в ротовую полость для добавочного пережёвывания[237].
Желудок
Желудок у большинства млекопитающих является простым (однокамерным). У однопроходных он имеет форму простого мешка и лишён пищеварительных желёз, а у сумчатых, даманов, насекомоядных, хищных, непарнокопытных, грызунообразных, приматов желудок напоминает по форме реторту, причём в эпителии его стенок находятся железистые клетки, выделяющие желудочный сок[238][239].
В состав желудочного сока входит, прежде всего, соляная кислота (HCl). Она гидролизует жиры и крахмал, кроме того, создаваемая кислотой низкая величина pH в желудке способствует обеззараживанию пищи и активирует протеолитические ферменты, входящие в состав желудочного сока — пепсины, действующие как эндопептидазы, то есть направленные на расщепление пептидных связей внутри полипептидной цепи. Кроме пепсинов, в состав желудочного сока могут входить также липазы. Липазы и пепсиногены (предшественники пепсинов, из которых последние отщепляются при низких значениях pH) секретируются главными клетками стенки желудка, а HCl выделяется париетальными клетками[240]. Помимо вышеперечисленных компонентов, в желудочном соке содержатся слизь и бикарбонат (HCO3-), защищающие слизистую оболочку желудка от твёрдых компонентов пищи и действия собственных секретов[241].
Однако, у некоторых млекопитающих, особенно у питающихся грубыми и трудно перевариваемыми растительными кормами, развился сложный (многокамерный) желудок, состоящий из нескольких обособленных отделов. Наиболее сложно в морфологическом и физиологическом плане он устроен у жвачных, обладающих четырёхкамерным желудком: в первых трёх его отделах — рубце, сетке, книжке[242] — пища подвергается брожению с участием симбионтов — бактерий и простейших, а пищеварительные железы имеются лишь в стенках четвёртого отдела (сычуг), в котором пища и подвергается воздействию желудочного сока[238].
У большинства других парнокопытных желудок трёхкамерный, только у свиней желудок простой. С питанием грубой растительной пищей связано наличие сложного желудка также у сирен, обладающих двухкамерным желудком, и трёхпалых ленивцев, имеющих пятикамерный желудок[243].
Несколько иное устройство имеет трёхкамерный желудок китообразных: их зубы не способны размельчать пищу, а желудок обеспечивает её механическую обработку при перистальтических движениях стенок[238]. У лишённых зубов муравьедов и панголинов желудок состоит из двух отделов: железистого и мускулистого. Последний и выполняет функции зубов по перетиранию пищи, причём у муравьедов такое перетирание обеспечивают очень твёрдые складки стенок данного отдела желудка, а у панголинов инструментом перетирания служит особая складка, которая вооружена роговыми зубами и вдаётся в полость мускулистого отдела, выстланную ороговевшим эпителием[244][245].
Кишечник
Кишечник млекопитающего подразделяют на тонкий и толстый. К тонкому кишечнику относят двенадцатиперстная кишка, тощая кишка и подвздошная кишка; к толстому — слепая кишка, ободочная кишка и прямая кишка[246][247].
В тонком кишечнике пища переваривается под влиянием пищеварительных соков. Они выделяются железами стенок кишки, а также печенью и поджелудочной железой, протоки которых впадают в начальный отдел тонкого кишечника[248].
Панкреатический сок, выделяемый поджелудочной железой, имеет щелочную реакцию и содержит ряд ферментов, секретируемых клетками поджелудочной железы. К числу этих ферментов относят:
- протеолитические ферменты группы эндопептидаз, высвобождаемые, как и пепсин, сначала в неактивной форме — трипсин и химотрипсин;
- карбоксипептидаза — протеолитический фермент группы эктопептидаз, отщепляющий отдельные аминокислоты от конца полипептида после того, как тот подвергся действию пепсина, трипсина и химотрипсина;
- панкреатическая α-амилаза, гидролизующая 1,4—гликозидные связи в полисахаридах, под её действием крахмал и гликоген расщепляются до мальтозы, мальтотриозы и α-декстрина;
- липазы — группа ферментов, расщепляющих жиры на жирные кислоты и моноглицериды. Их активность зависит от присутствия коферментов — колипаз;
- рибонуклеазы и дезоксирибонуклеазы, воздействующие на РНК и ДНК, соответственно;
- фосфолипаза A2, селективно расщепляющая фосфолипиды[249];
- эластазы, расщепляющие соединительную ткань пищи[250][251].
Жёлчь — пищеварительный секрет печени, её основными функциями являются облегчение всасывания продуктов расщепления жиров в тонкой кишке и эмульгирование жиров, нейтрализация кислого желудочного сока, поступающего в двенадцатиперстную кишку вместе с полупереваренной пищей (химусом), а также активизация моторики тонкого кишка, стимуляция продукции слизи и гастроинтенсинальных гормонов. В состав жёлчи входят: электролиты, соли жёлчных кислот, обеспечивающие эмульгирование, холестерин, лецитин (фосфатидилхолин), диглюкуронид билирубина, стероидные гормоны, лекарства и др[252]. Накопление и хранение жёлчи происходит в жёлчном пузыре, откуда она поступает в двенадцатиперстную кишку[253][254].
Через стенки тонкого кишечника основные питательные вещества всасываются в кровь, а остатки непереваренной пищи поступают в толстую кишку[248]. Длинноцепочечные жирные кислоты, нерастворимые в воде, всасываются не в кровь, а в кишечную лимфу, потом ещё раз проходят через печень и, наконец, попадают в большой круг кровообращения[255].
На границе тонкого и толстого кишечников расположен илеоцекальный клапан, не дающий формирующимся в толстом кишечнике каловым массам забрасываться назад в тонкий кишечник[256]. В толстом кишечнике трудно перевариваемые частицы пищи подвергаются бродильным процессам с участием эндосимбионтов — бактерий, грибков и простейших. Для видов, кормящихся грубой растительной пищей, особо важную роль в этих процессах играет слепая кишка, нередко несущая червеобразный отросток — аппендикс. У коалы, зайцеобразных, бобров, ондатры она достигает особо значительных относительных размеров (а даман имеет даже три слепые кишки[257])[248][258].
Для эффективного усваивания переработанной в толстом кишечнике пищи зайцеобразные прибегают к копрофагии: поедают свой кал и подвергают его вторичному перевариванию в желудке. Такое поведение — характерный признак отряда зайцеобразных[259]; впрочем, копрофагию и вторичное переваривание практикуют и некоторые грызуны (в частности, бобры, хомяки, шиншиллы, морские свинки, капибара)[260][261].
Для большинства млекопитающих характерно наличие в стенках слепой кишки большого количества лимфатической ткани, что делает её важным органом иммунной системы. У человека слепая кишка особой роли в пищеварении не играет[248][262], а у плотоядных видов она развита слабо или вообще отсутствует[263].
В ободочной кишке каловые массы обезвоживаются, скапливаются в прямой кишке и затем удаляются из организма через анальное отверстие[262].
Общая длина кишечника у млекопитающих очень сильно варьирует, отвечая составу их пищи. Обычно его относительная длина значительно больше у растительноядных видов. Так, у летучих мышей кишечник длиннее тела в 1,5—4 раза, у насекомоядных — в 2,5—4,2 раза, у грызунов — от 5,0 (полуденная песчанка) до 11,5 (морская свинка), у волка — в 6,5 раз, у лошади — в 12, у овцы — в 29 раз[263][264].
Выделительная система и осморегуляция

- 1 — корковый слой,
- 2 — мозговой слой,
- 3 — почечная артерия,
- 4 — почечная вена,
- 5 — мочеточник,
- 6 — нефроны,
- 7 — приносящая артериола,
- 8 — сосудистый клубочек,
- 9 — капсула Шумлянского — Боумена,
- 10 — почечные канальцы и петля Генле,
- 11 — выносящая артериола,
- 12 — околоканальцевые капилляры.
Система органов, формирующих, накапливающих и выделяющих мочу, млекопитающих состоит из пары почек, двух мочеточников, мочевого пузыря и мочеиспускательного канала[265].
Почки млекопитающих, как и у других наземных позвоночных, тазовые (метанефрические). Почки осуществляют водно-солевой обмен организма и выводят из него токсичные продукты обмена веществ. Конечными продуктами белкового обмена являются мочевина и мочевая кислота. Мочевина образуется в печени из аммиака, а мочевая кислота — продукт распада пуринов[266]. При этом у млекопитающих мочевина по содержанию в моче резко преобладает над мочевой кислотой (у рептилий и птиц соотношение обратное)[267].
Почки млекопитающих имеют бобовидную форму и располагаются в поясничной области, по бокам позвоночника. У китообразных и некоторых хищных почки состоят из нескольких долек. Каждая почка состоит из коркового и мозгового вещества, обе эти области сообщаются с почечными артериями и венами[268]. Моча, образующаяся в почке при фильтрации плазмы крови, поступает в почечную лоханку, откуда по мочеточнику небольшими порциями попадает в мочевой пузырь, где накапливается; затем она по мочеиспускательному каналу выходит наружу[269].
Паренхима почки представлена эпителиальными почечными канальцами, которые совместно с кровеносными капиллярами образуют нефроны. Нефрон — структурно-функциональная единица почки; в каждой почке млекопитающего содержатся тысячи нефронов (у домовой мыши — около 5 тысяч, у человека — около миллиона)[270][271]. Нефрон состоит из почечного тельца и выделительной трубочки; в свою очередь, в состав почечного тельца входят сосудистый клубочек с сетью капилляров и окружающая его двуслойная капсула Шумлянского — Боумена. Под давлением крови некоторое количество плазмы попадает из капилляров в пространство капсулы; образовавшийся фильтрат называется первичной мочой, которая отличается от плазмы крови отсутствием форменных элементов крови, а также малым количеством белков-анионов. В нефроне она подвергается дальнейшей фильтрации и движется по отделам выделительной трубочки — проксимальному канальцу, петле Генле и дистальному извитому канальцу. Нефрон открывается в собирательную трубочку; по ней моча выходит из нефрона, а потом — в конечном счёте — переходит в почечную лоханку[272][273].

В проксимальном канальце происходит реабсорбция ионов натрия, калия, хлора, гидрокарбонат-иона, воды, полное всасывание глюкозы, белков, витаминов, аминокислот. Клетки эпителия выделительной трубочки выделяют в фильтрат ионы аммония и водорода, регулируя тем самым pH организма; в результате азот находится в моче млекопитающих не только в виде мочевины, но и в виде ионов аммония. В нисходящей части петли Генле вода реабсорбируется через особые каналы, образованные белком аквапорином, однако каналов для ионов и других веществ здесь почти нет. В восходящей части происходит реабсорбция NaCl. Дистальный извитой каналец играет ключевую роль в регуляции концентрации ионов в организме за счёт изменения количеств секретируемых в мочу ионов К+ и реабсорбируемых NaCl. Наконец, в собирательной трубочке происходит всасывание воды, опосредованное аквапоринами[272][273].
Количество реабсорбируемой воды можно изменять, уменьшая или увеличивая число аквапоринов, и тем самым изменять осмолярность мочи и её количество. Осмолярность мочи также можно регулировать, изменяя осмолярность тканевой жидкости и тем самым изменяя параметры реабсорбции ионов. Эти механизмы дают возможность получать концентрированную мочу в условиях недостатка влаги и разбавленную — при избытке воды[272][273]. Вообще, количество мочи и её осмолярность находятся под контролем ряда гормонов, в частности, вазопрессина, ренин-ангиотензин-альдостероновой системы и предсердного натрийуретического пептида[274].
Среди позвоночных способностью транспортировать соли сквозь стенки почечного канальца обладают только млекопитающие и птицы. У последних данная способность выражена слабее, и осмотическое давление их мочи может превышать такой показатель для плазмы крови лишь вдвое, а у млекопитающих — в 25 раз[174].
Выделяют три типа нефронов: суперфициальные, расположенные поверхностные в коре и имеющие короткую петлю Генле (примерно 15 % всех нефронов); интракортикальные, играющие основную роль в ультрафильтрации мочи (70 %); юкстамедуллярные, выполняющие основную роль в процессах концентрирования и разведения мочи (15 %)[271][275]. Длина петли Генле у юкстамедуллярных нефронов в значительной мере определяется условиями обитания того или иного вида. Так, у нефронов млекопитающих, живущих в засушливых районах, петля Генле длинная, глубоко уходящая в мозговое вещество почки. Длинная петля Генле поддерживает высокий осмотический градиент в почках, в результате чего образуется гиперосмотичная моча. Напротив, у нефронов водных млекопитающих, не испытывающих проблем с дегидратацией, петля Генле значительно короче, что ограничивает их возможности по концентрированию мочи. Наземные млекопитающие, населяющие умеренно влажные районы, имеют нефроны со средней длиной петли Генле, и они могут выделять мочу промежуточной степени концентрирования. Таким образом, наличие у млекопитающих юкстамедуллярных нефронов можно считать приспособлением к наземному образу жизни[276].
Помимо почек, определённую роль в выделении играют потовые железы и лёгкие, а также печень. Наиболее важная функция последней — образовании мочевины из избытка аминокислот. Кроме того, в составе жёлчи печень выводит жёлчные пигменты, образующиеся при разрушении гемоглобина старых эритроцитов. Эти пигменты выходят из организма в составе кала, придавая ему характерный цвет[277].
Эндокринная система

Эндокринную систему составляют железы внутренней секреции, регулирующие работу различных органов тела. Эти железы не имеют выводных протоков и выделяют вырабатываемые ими гормоны в кровь (реже — в лимфу)[34]. У млекопитающих эндокринная система принципиально сходна с подобными системами других позвоночных, отличаясь не столько составом или числом гормонов, сколько усилением их активности и специализации[278].
Тем не менее, некоторые гормоны у млекопитающих имеют другое действие, чем у остальных позвоночных. Так, меланоцитстимулирущий гормон (МСГ) у земноводных, рыб и рептилий регулирует цвет кожи, контролируя распределение пигмента в клетках кожи, называемых меланоцитами. У млекопитающих МСГ не только влияет на распределение пигмента в коже, но действует на обмен веществ и отвечает за чувство голода. Другим примером может служить пролактин, который у пресноводных рыб регулирует водно-солевой обмен, у земноводных — тормозит метаморфоз, у птиц — регулирует метаболизм жиров и репродуктивные функции, а у млекопитающих стимулирует рост млечных желёз и образование молока[279].
К числу важнейших эндокринных желёз относят:
- Гипоталамус — центральная структура регуляции эндокринных функций. Его нейроны синтезируют и секретируют специальные регуляторные пептиды — нейрогормоны, транспортируемые в структуры мозга, ликвор или гипофиз. Эти нейрогормоны могут преимущественно действовать на висцеральные органы (окситоцин, вазопрессин), на функции нервной системы (эндорфины, энкефалины, нейротензин, вазопрессин, ангиотензин и др.) или же на железистые клетки аденогипофиза (рилизинг-гормоны: либерины — усиливают секрецию, статины — подавляют)[280].
- Гипофиз. Он имеет три отдела, задний из которых — нейрогипофиз — секретирует вазопрессин и окситоцин (первый в основном регулирует водно-солевой обмен, второй — сокращения гладких мышц при родах, лактации), а передний — аденогипофиз — выделяет несколько гормонов, причём функции его железистых клеток находятся под контролем гипоталамуса, осуществляемым через вышеупомянутые нейрогормоны. В аденогипофизе различают пять групп хромафильных клеток, вырабатывающих следующие гормоны: соматотропин (или гормон роста), пролактин, тиреотропин, гонадотропины (фолликулостимулирующий и лютеинизирующий), тиреотропин (или адренокортикотропный гормон). Кортикотропин, гонадотропины и тиреотропин регулируют соответственно кору надпочечников, половые железы и щитовидную железу, а соматотропин и пролактин действуют на различные ткани-мишени[281].
- Эпифиз. Вырабатываемый им гормон — мелатонин — участвует в регуляции циркадных ритмов[282].
- Щитовидная железа. Важнейший из её гормонов — тироксин — регулирует развитие организма и обмен веществ, другой гормон — кальцитонин — участвует в кальциевом обмене (снижает уровень кальция в крови)[283].
- Паращитовидные железы. Вырабатываемый ими паратгормон, прежде всего, регулирует кальциевый обмен и является антагонистом кальцитонина[284].
- Надпочечники. Их мозговое вещество отвечает за выработку катехоламинов — адреналина и норадреналина, влияющих на кровообращение, углеводный обмен и работу нервной системы. В корковом слое надпочечников синтезируются стероидные гормоны: минералокортикоид альдостерон регулирует водно-солевой обмен, глюкокортикоиды имеют широкий спектр действия на метаболизм и половые стероиды[285].

Некоторые органы выполняет одновременно несколько важных эндокринных и неэндокринных функций. Так, поджелудочная железа, выделяющая пищеварительные ферменты, секретирует также и ряд гормонов. Такую секрецию осуществляют островки Лангерганса — скопления клеток нескольких типов, которые выделяют следующие гормоны (участвующие, прежде всего, в регуляции углеводного обмена): инсулин (β-клетки), глюкагон (α-клетки), соматостатин (δ-клетки), панкреатический полипептид (РР-клетки), грелин (ε-клетки)[286]. При этом соотношение келточных составляющих островков Лангерганса различна у разных видов млекопитающих. Так, у грызунов почти вся центральнаю часть островка занята β-клетками, а остальные клетки оттеснены к периферии; у человека же α- и β-клетки равномерно располагаются по всему островку[287]. Печень, выделяющая жёлчь, тоже секретирует ряд гормонов: ангиотензин, тромбопоэтин, гепцидин, бетатрофин, а также инсулиноподобный фактор роста 1[288].
Половые железы — семенники и яичники — также сочетают эндокринные и неэндокринные функции: в них вырабатываются гаметы — сперматозоиды и яйцеклетки, а также образуются половые гормоны, важнейшими из которых являются тестостерон и эстрогены соответственно. Другой важный женский гормон, необходимый для овуляции — прогестерон — выделяется в основном клетками жёлтого тела[289].
У млекопитающих множество гормонов образуется в органах и тканях, не относящихся к эндокринным железам, но содержащих эндокринные клетки[290]. К числу таких органов и тканей относят следующие.
- Желудочно-кишечный тракт. Клетки слизистой оболочки желудка и двенадцатиперстной кишки секретируют множество пептидов, оказывающих преимущественно местное действие[291]. Среди таких интестинальных гормонов: гастрин, вырабатываемый желудком, который стимулирует секрецию пепсина и активирует моторику желудка; холецистокинин, секретируемый преимущественно в двенадцатиперстной кишке, который активирует пищеварение в тонкой кишке[292]; грелин, вырабатываемый париетальными клетками желудка и усиливающий аппетит[291], и другие.
- Плацента. Кроме обеспечения питания и вывода метаболитов от плода, она также выделяет ряд гормонов — прежде всего хорионический гонадотропин, тормозящий развитие фолликулов в яичниках матери и поддерживающий синтез прогестерона и эстрогена в жёлтом теле до тех пор, пока плацента не будет способна сама синтезировать их достаточные количества. Помимо хорионического гонадотропина, эстрогена и прогестерона плацента синтезирует кортикотропин-рилизинг-гормон (кортиколиберин), плацентарный лактоген и проопиомеланокортин[293].
- Тимус (вилочковая железа). Обеспечивает продукцию специфических Т-лимфоцитов и выделяет также гормоны группы тимозинов и тимопоэтин, которые управляют процессом созревания T-лимфоцитов[294], а также регулируют процессы синтеза клеточных рецепторов к гормонам и нейромедиаторам, стимулируют разрушение ацетилхолина в нервно-мышечных синапсах и участвуют в регуляции других процессов, важных для всего организма[295].
- Почки. Некоторые их клетки секретируют гормоны кальцитриол (третий кальцийрегулирующий гормон), ренин (начальное звено ренин-ангиотензин-альдостероновой системы) и эритропоэтин[295] (стимулирует образование эритроцитов в красном костном мозге (эритропоэз)[296]).
- Сердце. Кардиомиоциты сердца выделяют предсердный натрийуретический пептид — мощный вазодилататор и антагонист ренин-ангиотензин-альдостероновой системы[297]. В предсердиях в малых количествах образуются также соматостатин и ангиотензин-II[291].
- Жировая ткань. Её клетки (адипоциты) секретируют гормон лептин, подавляющий аппетит и выступающий тем самым антагонистом грелина[291]).
Гормоны млекопитающих представлены разными классами органических соединений. Среди них есть пептиды (окситоцин, вазопрессин, адренокортикотропный гормон, паратгормон, кальцитонин), белки (гормон роста, пролактин, инсулин, глюкагон), гликопротеины (фолликулостимулирующий гормон, лютеинизирующий гормон, тиреотропный гормон), производные аминокислот (трийодтиронин, тироксин, адреналин, норадреналин, мелатонин) и стероиды (глюкокортикоиды, минералокортикоиды, андрогены, эстрогены, прогестерон)[282].
Репродуктивная система
Половые органы

Половые железы самцов млекопитающих — семенники — имеют характерную овальную форму. У однопроходных, неполнозубых, броненосцев, хоботных и китообразных они в течение всей жизни животного остаются в полости тела, но у большинства других млекопитающих со временем семенники опускаются в расположенный снаружи кожистый мешочек — мошонку. Через семявыводящие каналы и парный семяпровод вырабатываемые семенниками сперматозоиды поступают в мочеиспускательный канал, располагающийся у самца на нижней части пениса. В этот канал свой жидкий секрет выделяют предстательная железа и куперовы железы. В результате формируется сперма. Во время полового акта она попадает во влагалище (у однопроходных — в клоаку) самки[298].

Имеющиеся в пенисе пещеристые тела при половом возбуждении наполняются кровью, благодаря чему он становится упругим и увеличивается в объёме. Дополнительную прочность пенису придаёт имеющаяся у ряда млекопитающих (сумчатые, китообразные, хищные, некоторые рукокрылые, грызуны и приматы) в конечной его части непарная кость — бакулюм. У сумчатых пенис часто бывает раздвоен в соответствии с наличием двух влагалищ у самок[299].
Половые железы самки млекопитающих — яичники — всегда лежат в полости тела; туда же своими воронками открываются парные яйцеводы. Созревшая яйцеклетка (у однопроходных она достаточно велика: 2,5—4,0 мм, а у сумчатых и плацентарных микроскопически мала: 0,2—0,4 и 0,05—0,2 мм соответственно) во время овуляции через полость тела попадает в фаллопиеву трубу — верхний извитой отдел яйцевода. В фаллопиевой трубе (у обыкновенного тенрека — в яичнике) и происходит оплодотворение; образовавшаяся зигота тут же начинает дробиться и передвигается в матку, где и происходит развитие эмбриона[300][301].
Репродуктивные циклы самок
Овариальный цикл
Если у самцов млекопитающих гаметы образуются постоянно после наступления полового созревания, то у самок гаметы созревают циклически, и вместе с ними циклически изменяются уровни женских половых гормонов, а также происходят некоторые циклические изменения в женских репродуктивных органах. Циклический последовательный ряд физиологических процессов созревания яйцеклетки и фолликула получил название овариального, или яичникового, цикла[302].
Менструальный цикл

Менструальным, или маточным, циклом называют циклические изменения эндометрия матки в ответ на циклическую секрецию женских половых стероидов. Менструальный цикл характерен для человека, а также некоторых приматов[303]. Менструальный цикл включает три фазы: менструальную, пролиферативную и секреторную[304].
Менструальная фаза представляет собой кровотечение из полости матки в результате отторжения слоя эндометрия, которое происходит в конце овариального цикла, если не было оплодотворения яйцеклетки. Пролиферативная фаза совпадает с фолликулярной и овуляторной фазами овариального цикла. В ходе этой фазы под действием эстрогенов происходит клеточная пролиферация тканей органов женской половой системы, происходящие в ходе неё изменения направлены на облегчение возможного в будущем движения сперматозоидов к яйцеклетке; кроме того, увеличивается число рецепторов к прогестерону. Секреторная фаза начинается непосредственно после овуляции, когда возрастает уровень прогестерона в крови за счёт выделения этого гормона жёлтым телом. Прогестерон и эстрадиол жёлтого тела регулируют развитие слизистой оболочки и кровеносных сосудов в эндометрии, а также усиливают секрецию слизи и синтез веществ, необходимых для поддержания метаболизма оплодотворённой яйцеклетки и способствуют её внедрению в оболочку матки. Если оплодотворения не происходит, то из-за лютеолиза уровень прогестерона и эстрадиола падает, тонус гладких мышц артерий эндометрия увеличивается, что вызывает его ишемию, некроз и менструальное кровотечение[305].
У млекопитающих, имеющих менструальный цикл, самки способны к половому поведению в любой момент цикла. Однако впоследствии (у человека — в возрасте 46—54 лет, после приблизительно 500 циклов) у них наступает прекращение менструаций — менопауза. При менопаузе яичники становятся нечувствительными к фоллитропину и лютеотропину[303].
Эстральный цикл
У большей части самок млекопитающих, однако, менструального цикла нет, хотя перед овуляцией эндометрий у них также утолщается. У самок этих млекопитающих в отсутствие оплодотворения происходит только реабсорбция эндометрия и отсутствует маточное кровотечение. У них нет менопаузы, кроме того, они способны к проявлению половой активности и к спариванию не в любой момент цикла, как у самок, имеющих менструальный цикл, а лишь в конкретный период, называемый эструсом, или течкой. Изменения в эндометрии и в этом случае происходит циклически: имеет место эстральный цикл[303][306].
Продолжительность и частота эстрального цикла у различных млекопитающих существенно различаются. Выделяют моноэстральные виды, для которых характерен лишь один эстральный цикл в течение года, и полиэстральные виды, имеющие в течение года несколько эстральных циклов. К моноэстральным видам относятся однопроходные, часть сумчатых, кроты, многие хищные (медведи, волки, лисицы и др.), некоторые грызуны (бобры, сурки), косули. У них эструс обычно приурочен к определённому времени года, а бо́льшую часть года составляет анэструс — период, когда самка к спариванию неспособна[306].
К полиэстральным видам относятся многие грызуны и копытные, львы, гиены и др. Длительность эстрального цикла у таких млекопитающих обычно уменьшается с уменьшением размеров животного: так, у африканских слонов цикл длится 16 недель, у крыс — 5 дней, у сирийского хомячка — всего 4 дня[303][307]. У коров длина эстрального цикла составляет в среднем 21 день[308] и продолжительность эструса составляет 18—24 часа, а у лошадей эструс длится 4—7 дней при длине эстрального цикла 19—25 дней[309].
Эстральный цикл включает 4 фазы: проэструс, эструс, метэструс, диэструс. Во время проэструса начинается рост одного или нескольких фолликулов, усиливается секреция эстрогенов, происходит утолщение эндометрия. Фазу эструса характеризуют наивысший уровень секреции эстрогенов и повышенная сексуальная восприимчивость самки, нередко проявляющаяся в активном поиске сближения с самцом. Обычно эструс сопровождается овуляцией, хотя у некоторых насекомоядных, хищных, грызунов и большинства зайцеобразных овуляция происходит только после спаривания. Если оплодотворения и последующей имплантации зародыша не состоялось, возбуждение сменяется торможением: наступает фаза метэструса, во время которой секреция эстрогенов падает, а секреция прогестерона, напротив, достигает пика. Заключительной фазе цикла — диэструсу — соответствует низкий уровень секреции всех гормонов, производимых яичниками. У полиэстральных видов относительно короткий диэструс сменяется новым проэструсом, а у моноэстральных видов наступает длительный анэструс[310].
Соотношения между длительностями фаз эстрального цикла видоспецифичны. Так, у серой крысы проэструс и эструс длятся по 12 ч, метэструс — 21 ч, диэструс — 57 ч; у домашней свиньи соответственно — 3, 2—3, 3 и 11—13 дней[311][312].
Определение пола

Млекопитающим присущ гетерогаметный способ определения пола: имеются диморфные половые хромосомы X и Y, причём геном самцов содержит и хромосому X, и хромосому Y, а геном самок — две хромосомы X. У плацентарных млекопитающих пол определяется геном SRY на дистальном участке короткого плеча Y-хромосомы; данный ген проявляет кратковременную активность в онтогенезе, инициируя работу семенной железы. Если данного гена нет или работу его подавить, то гонада развивается в яичник, секретирующий гормоны, которые в дальнейшем определяют женский тип развития организма[313].
Резко отличен способ определения пола у утконоса: у него пол задаётся не одной, а пятью парами половых хромосом (самец имеет набор хромосом XYXYXYXYXY, а самка — XXXXXXXXXX)[314].
В 1990-х гг. выявлены особенности определения пола у некоторых грызунов. У одного из видов слепушонок оба пола имеют набор половых хромосом XO, у другого вида — XX, а у третьего из изученных видов механизм определения пола — обычный для млекопитающих[315]. У рюкюских колючих мышей также отсутствует Y-хромосома, а значит, и ген SRY[316].
Вынашивание плода
Матка представляет собой нижний расширенный отдел яйцевода. У однопроходных и сумчатых матки полностью разделены. У плацентарных различают[317][318]:
- двойную матку: каждая матка открывается во влагалище самостоятельным отверстием (многие грызунообразные, трубкозубые, даманы, хоботные);
- двураздельную матку: обе матки срослись концами и имеют общее выводное отверстие (ряд грызунов, некоторые хищные и копытные);
- двурогую матку: матки срослись на половину длины (насекомоядные, ряд рукокрылых, хищных, копытных, китообразных и низшие приматы);
- простую матку: матки полностью слились в единую матку (ксенартры, некоторые рукокрылые, высшие приматы).
У однопроходных плацента не образуется, а беременность длится 14 суток у утконоса и от 16 до 27 дней у ехидновых и завершается откладыванием яиц размером 1—2 см в ороговевшей скорлупе, содержащих более чем наполовину развившийся эмбрион. Затем самка утконоса высиживает яйца в норе в течение 9—10 суток, а самка ехидны вынашивает их 10—11 дней в кожистой сумке. Вылупившись, детёныш остаётся в сумке ещё довольно долго[319].

У сумчатых во время беременности уже возникает плацента — специфический орган связи плода и материнского организма, образующийся путём срастания наружной оболочки зародыша (хориона) и разрыхлённого эпителия матки. Этот орган обеспечивает снабжение зародыша кислородом и питательными веществами и выведение углекислоты и продуктов обмена[320]. Однако плацента сумчатых примитивная — хориовителлиновая, связывающая зародыш и организм матери через кровеносные сосуды желточного мешка (у бандикутов плацента — хориоаллантоисная, то есть обеспечивает связь зародыша с организмом матери через кровеносные сосуды аллантоиса, но лишена ворсинок, характерных для плацентарных). В результате период внутриутробной жизни у сумчатых короток, составляя от 8 до 42 дней (у опоссумов — 12—13, коалы — 30—35, гигантского кенгуру — около 36 дней), а детёныш рождается сильно недоношенным (размеры детёныша двухметрового гигантского кенгуру не превышают 25 мм, а весит он менее 1 г). После рождения детёныш столь слаб, что неделями висит, прикрепившись к соску, и не в состоянии даже сосать; мать сама периодически впрыскивает, сокращая мышцы млечного поля, молоко ему в рот[321][322].

Намного более совершенно устройство хориоаллантоисной плаценты у плацентарных. У них эмбрион имеет уникальный наружный слой — трофобласт, который формирует многочисленные ворсинки хориона, глубоко прорастающие в эпителий матки. При этом клетки трофобласта создают между материнской и эмбриональной тканями активный барьер, препятствующий отторжению плода из-за несовместимости тканей; это делает возможным существенное продление периода внутриутробного развития, в течение которого плод надёжно защищён от неблагоприятных условий внешней среды[321].
Сроки вынашивания плода и степень развития новорождённых у различных видов плацентарных зависят от размеров животного: у сирийского хомячка она составляет 16 дней, а у слонов — почти два года (у индийского слона 623—729 дней, у саванного слона — 640—673 дней)[323][324].
Однако, ещё сильнее сроки беременности зависят от условий жизни детёнышей после родов, и в этом плане говорят о имматуронатных (незрелорождающих) и матуронатных (зрелорождающих) зверях[325]. К первой группе относят прежде всего животных, имеющих хорошо устроенные убежища. У них беременность короче, а новорождённые появляются на свет беспомощными, слепыми, часто голыми; так, у домовой мыши беременность длится 19 дней, у кролика — 28—32, белки — 35—40, лисицы — 49—58, кошки — 58—65, волка — 60—68. Звери же, не имеющие таких убежищ, обычно относятся ко второй группе: у них беременность длится дольше, а детёныши рождаются зрячими, покрытыми шерстью и способными к передвижению; таковы, например, детёныши зайца-беляка и нутрии (у этих видов беременность длится соответственно 49—51 и 128—133 дня)[326][327]. Особенно продолжительна беременность у животных, чьи детёныши способны уже через несколько часов после рождения следовать за матерью (копытные, китообразные): свинья — 112—115 дней, лось — 240—250, корова — 279—292, лошадь — 330—342, синий кит — 330—360, жираф — 420—450[326][328].
К значительному удлинению сроков беременности приводит эмбриональная диапауза (задержка имплантации эмбриона в стенку матки). Оно обнаружено почти у ста видов, относящихся к 7 отрядам млекопитающих (в основном — к хищным и двурезцовым)[329]. У канадской выдры из-за задержки имплантации срок вынашивания плода возрастает до 10—12 месяцев (в то время как у обыкновенной выдры беременность длится в среднем 63 дня)[330]. У американской норки имплантация задерживается на срок от 8 до 45 дней, вследствие чего продолжительность беременности варьирует от 40 до 75 дней. У барсука задержка имплантации, достигающая 10 месяцев, позволяет вне зависимости от момента спаривания приурочить имплантацию к февралю, а рождение детёнышей — к концу марта или началу апреля. Несмотря на столь длительную беременность, все упомянутые виды — имматуронатные[331].
Половой диморфизм

Почти у всех видов млекопитающих наблюдается половой диморфизм — различия между самцами и самками одного и того же биологического вида, исключая различия в строении половых органов. Некоторые примеры полового диморфизма у млекопитающих приведены ниже.
- Размер тела. Как правило, у млекопитающих взрослые самцы крупнее самок (например, у саванного слона масса самца в среднем примерно в два раза превосходит массу самки[332], а самец северного морского котика весит в 4,5 раза больше, чем самка[333]). Однако известны исключения: например, у синего кита самка тяжелее самца и на 1—2 м длиннее[334].
- Окрас меха. У млекопитающих половой дихроматизм выражен в меньшей мере, чем у птиц, у которых самцы, как правило, окрашены более ярко, чем самки. У тех видов млекопитающих, у которых присутствует половой дихроматизм, самки обычно предпочитают более ярко окрашенных самцов. Это можно объяснить, во-первых, тем, что более яркий окрас свидетельствует о хорошем состоянии здоровья самца и его способности дать здоровое потомство, и, во-вторых, тем, что сам факт доживания ярко окрашенного самца до репродуктивного периода говорит о его способности избежать хищников[335].
- Болезни. Некоторые болезни распространены исключительно или преимущественно среди особей только одного пола одного биологического вида. Это касается как генетически сцеплённых с полом болезней (в частности, гемофилия у человека), так и болезней, затрагивающих органы, присущие только самкам или самцам или развитых в полной мере только у особей одного пола (например, болезней матки и млечных желёз)[336].
Примечания
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 369—370.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 309—311.
- ↑ 3,0 3,1 Proksch E., Brandner J. M., Jensen J. M. The skin: an indispensable barrier // Experimental dermatology. — 2008. — Vol. 17, № 12. — P. 1063—1072. — doi:10.1111/j.1600-0625.2008.00786.x. — PMID 19043850.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 65, 293, 371.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 371.
- ↑ Khanna D. R., Yadav P. R. . Biology of Mammals. — New Delhi: Discovery Publishing House, 2005. — viii + 445 p. — ISBN 81-7141-934-8. — P. 97.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 24—25, 312.
- ↑ 8,0 8,1 8,2 Дзержинский, Васильев, Малахов, 2014, с. 372—373.
- ↑ Кэрролл, т. 2, 1993, с. 236, 238.
- ↑ Константинов, Шаталова, 2004, с. 364.
- ↑ Медников, 1994, с. 343.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 313—314.
- ↑ 13,0 13,1 13,2 Константинов, Шаталова, 2004, с. 365.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 314.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 314—315.
- ↑ Antlers // Hall B. K. Bones and Cartilage: Developmental and Evolutionary Skeletal Biology. — San Diego, London: Elsevier Academic Press, 2005. — xxviii + 760 p. — ISBN 0-12-319060-6. — P. 103—114.
- ↑ Кэрролл, т. 3, 1993, с. 87.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 373.
- ↑ Константинов, Шаталова, 2004, с. 366.
- ↑ Разнообразие млекопитающих, ч. III, 2004, с. 811.
- ↑ 21,0 21,1 Медников, 1994, с. 371.
- ↑ 22,0 22,1 Кэрролл, т. 2, 1993, с. 236.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 118, 315.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 293, 375.
- ↑ Константинов, Шаталова, 2004, с. 367.
- ↑ Walcott B. The Lacrimal Gland and Its Veil of Tears // News in Physiological Sciences. — 1998. — Vol. 13. — P. 97—103. — PMID 11390770.
- ↑ 27,0 27,1 Константинов, Наумов, Шаталова, 2012, с. 315.
- ↑ Francis C. M., Anthony E. L. P., Brunton J. A., Kunz T. H. Lactation in male fruit bats (англ.) // Nature. — 1994. — Vol. 367, № 6465. — P. 691–692. — doi:10.1038/367691a0..
- ↑ Kunz T. H., Hosken D. J. Male lactation: why, why not and is it care? // Trends in Ecology and Evolution. — 2009. — Vol. 24, № 2. — P. 80—85. — doi:10.1016/j.tree.2008.09.009. — PMID 19100649.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 315—316.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 375, 417.
- ↑ Разнообразие млекопитающих, ч. III, 2004, с. 682, 690, 722.
- ↑ Kielan-Jaworowska Z. . In Pursuit of Early Mammals. — Bloomington: Indiana University Press, 2013. — xvii + 272 p. — ISBN 978-0-253-00817-6. — P. 219—220.
- ↑ 34,0 34,1 Медников, 1994, с. 371—372.
- ↑ Барабаш-Никифоров, Формозов, 1963, с. 48.
- ↑ 36,0 36,1 Кэрролл, т. 2, 1993, с. 238.
- ↑ 37,0 37,1 Константинов, Шаталова, 2004, с. 371.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 303, 377.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 378—379.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 255, 379—380, 438.
- ↑ 41,0 41,1 Rubidge B. S., Sidor C. A. Evolutionary Patterns among Permo-Triassic Therapsids (англ.) // Annual Review of Ecology, Evolution, and Systematics. — Annual Reviews, 2001. — Vol. 32. — P. 449–480. — doi:10.1146/annurev.ecolsys.32.081501.114113. Архивировано 19 декабря 2012 года.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 255—256, 305, 379—380.
- ↑ Константинов, Шаталова, 2004, с. 372.
- ↑ The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea / Ed. by J. G. M. Thewissen. — New York: Plenum Press, 1998. — xii + 479 p. — (Advances in Vertebrate Paleobiology). — ISBN 0-306-45853-5. — P. 44, 47.
- ↑ Hautier L., Weisbecker V., Sánchez-Villagra M. R. e. a. Skeletal development in sloths and the evolution of mammalian vertebral patterning // Proceedings of the National Academy of Sciences of the United States of America. — 2010. — Vol. 107, № 44. — P. 18903—18908. — doi:10.1073/pnas.1010335107. — PMID 20956304.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 375, 377.
- ↑ Константинов, Шаталова, 2004, с. 369—370.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 317—318.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 320.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 380.
- ↑ 51,0 51,1 Медников, 1994, с. 348.
- ↑ Константинов, Шаталова, 2004, с. 373.
- ↑ 53,0 53,1 Константинов, Шаталова, 2004, с. 373—374.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 379.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 192, 317.
- ↑ 56,0 56,1 Константинов, Шаталова, 2004, с. 374, 498.
- ↑ Зильбернагль, Деспопулос, 2013, с. 64.
- ↑ Daniel E. E., Tomita T., Tsuchida S., Watanabe M. . Sphincters: Normal Function — Changes in Diseases. — Boca Raton: CRC Press, 1992. — 464 p. — ISBN 0-8493-6748-4. — P. 3.
- ↑ 59,0 59,1 59,2 Ткаченко, 2009, с. 36.
- ↑ Зильбернагль, Деспопулос, 2013, с. 76.
- ↑ 61,0 61,1 Гистология, цитология и эмбриология, 2004, с. 310.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 385.
- ↑ 63,0 63,1 Нервная ткань — статья из Медицинской энциклопедии
- ↑ 64,0 64,1 Нервная ткань — статья из Большой советской энциклопедии.
- ↑ 65,0 65,1 Зильбернагль, Деспопулос, 2013, с. 48.
- ↑ Bean B. P. The action potential in mammalian central neurons // Nature reviews. Neuroscience. — 2007. — Vol. 8, № 6. — P. 451—465. — doi:10.1038/nrn2148. — PMID 17514198.
- ↑ Bennett M. V., Zukin R. S. Electrical coupling and neuronal synchronization in the Mammalian brain // Neuron. — 2004. — Vol. 41, № 4. — P. 495—511. — doi:10.1016/S0896-6273(04)00043-1. — PMID 14980200.
- ↑ Сидоров, 2008, с. 116.
- ↑ Campbell, 2011, p. 1057.
- ↑ 70,0 70,1 Жизнь животных, т. 6, 1971, с. 13.
- ↑ 71,0 71,1 Константинов, Шаталова, 2004, с. 387.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 246, 386.
- ↑ 73,0 73,1 Константинов, Наумов, Шаталова, 2012, с. 229, 333.
- ↑ 74,0 74,1 Дзержинский, Васильев, Малахов, 2014, с. 313, 386—387.
- ↑ Дзержинский, 2005, с. 263.
- ↑ Константинов, Шаталова, 2004, с. 387—389.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 333.
- ↑ Жизнь животных, т. 6, 1971, с. 283, 301, 554.
- ↑ Мак-Фарленд Д. . Поведение животных: психобиология, этология и эволюция. — М.: Мир, 1988. — 520 с. — ISBN 5-03-001230-2. — С. 460—461.
- ↑ Медников, 1994, с. 13—14.
- ↑ Бурлак С. А., Старостин С. А. . Сравнительно-историческое языкознание. — М.: Издат. центр «Академия», 2005. — 432 с. — ISBN 5-7695-1445-0. — С. 276—277, 281—282.
- ↑ Дзержинский, 2005, с. 263—264.
- ↑ Барабаш-Никифоров, Формозов, 1963, с. 59.
- ↑ 84,0 84,1 Барабаш-Никифоров, Формозов, 1963, с. 59—60.
- ↑ Дзержинский, 2005, с. 268—269.
- ↑ 86,0 86,1 Гистология, цитология и эмбриология, 2004, с. 340—341.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 391.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 336.
- ↑ Campbell, 2011, p. 1099.
- ↑ Гистология, цитология и эмбриология, 2004, с. 341—344.
- ↑ Джадд Д., Вышецки Г. . Цвет в науке и технике. — М.: Мир, 1978. — 592 с. — С. 16-18.
- ↑ Константинов, Шаталова, 2004, с. 209, 273, 391.
- ↑ Ткаченко, 2009, с. 389.
- ↑ Campbell, 2011, p. 1097.
- ↑ Bowmaker J. K. Evolution of colour vision in vertebrates // Eye (London, England). — 1998. — Vol. 12 (Pt 3b). — P. 541—547. — doi:10.1038/eye.1998.143. — PMID 9775215.
- ↑ Константинов, Шаталова, 2004, с. 391.
- ↑ Vaughan, Ryan, Czaplewski, 2011, p. 23.
- ↑ Лысов, Ипполитова, Максимов, Шевелёв, 2012, с. 79, 116.
- ↑ 99,0 99,1 Константинов, Шаталова, 2004, с. 390.
- ↑ 100,0 100,1 100,2 Дзержинский, Васильев, Малахов, 2014, с. 389.
- ↑ Nida Sophasarun. Experts Debunk Bats' Bad Rap. Online extra. National Geographic. Дата обращения: 30 апреля 2013. Архивировано 26 января 2018 года.
- ↑ Rachel Kaufman. Bats Crash More When They Rely on Vision (15 декабря 2010). Архивировано 1 апреля 2013 года. Дата обращения 30 апреля 2013.
- ↑ Гистология, цитология и эмбриология, 2004, с. 372—373.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 35, 335.
- ↑ Константинов, Шаталова, 2004, с. 389, 514.
- ↑ Гистология, цитология и эмбриология, 2004, с. 340—341, 372—373, 383.
- ↑ Зильбернагль, Деспопулос, 2013, с. 376.
- ↑ Наумов, Карташёв, 1979, с. 242.
- ↑ Campbell, 2011, p. 1093.
- ↑ 110,0 110,1 Ткаченко, 2009, с. 408—412.
- ↑ Campbell, 2011, p. 1091.
- ↑ Зильбернагль, Деспопулос, 2013, с. 376—377.
- ↑ Гистология, цитология и эмбриология, 2004, с. 375—377.
- ↑ Kelly M. C., Chen Ping. Shaping the mammalian auditory sensory organ by the planar cell polarity pathway // The International Journal of Developmental Biology. — 2007. — Vol. 51, № 6—7. — P. 535–547. — doi:10.1387/ijdb.072344mk. — PMID 4158833..
- ↑ Гистология, цитология и эмбриология, 2004, с. 377—380.
- ↑ Roy D. Development of Hearing in Vertebrates with Special Reference to Anuran Acoustic Communication // Journal of Biosciences. — 1994. — Vol. 19, № 5. — P. 629–644.
- ↑ Walsh S. A., Barrett P. M., Milner A. C., Manley G., Witmer L. M. Inner Ear Anatomy is a Proxy for Deducing Auditory Capability and Behaviour in Reptiles and Birds // Proc. of the Royal Society. B. — 2009. — Vol. 276, № 1660. — P. 1355–1360. — doi:10.1098/rspb.2008.1390. Архивировано 24 сентября 2015 года..
- ↑ 118,0 118,1 Зильбернагль, Деспопулос, 2013, с. 376—379.
- ↑ 119,0 119,1 Campbell, 2011, p. 1091—1092.
- ↑ Ткаченко, 2009, с. 406—408.
- ↑ Vaughan, Ryan, Czaplewski, 2011, p. 27.
- ↑ 122,0 122,1 Зильбернагль, Деспопулос, 2013, с. 352.
- ↑ Гистология, цитология и эмбриология, 2004, с. 363.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 335.
- ↑ Жизнь растений. Энциклопедия в 6 тт. Т. 2: Грибы / Гл. ред. А. А. Фёдоров. — М.: Просвещение, 1976. — 479 с. — С. 205.
- ↑ Meisami E., Bhatnagar K. P. Structure and diversity in mammalian accessory olfactory bulb // Microscopy Research and Technique. — 1998. — Vol. 43, no. 6. — P. 476—499. — doi:10.1002/(SICI)1097-0029(19981215)43:6<476::AID-JEMT2>3.0.CO;2-V. — PMID 9880163.
- ↑ Brennan P. A., Keverne E. B. . The vomeronasal organ // Handbook of Olfaction and Gustation. 2nd ed / Ed. by R. L. Doty. — New York: Marcel Dekker, 2003. — xiv + 1121 p. — (Neurological Disease and Therapy. Book 32). — ISBN 0-8247-0719-2. — P. 967—979.
- ↑ Swaney W. T., Keverne E. B. The evolution of pheromonal communication // Behavioural Brain Research. — 2009. — Vol. 200, no. 2. — P. 239—247. — doi:10.1016/j.bbr.2008.09.039. — PMID 18977248.
- ↑ Зильбернагль, Деспопулос, 2013, с. 352—353.
- ↑ 130,0 130,1 Ткаченко, 2009, с. 415.
- ↑ Воротников С. А. Информационные устройства робототехнических систем. — М.: Изд-во МГТУ им. Н. Э. Баумана, 2005. — 384 с. — ISBN 5-7038-2207-6. — С. 307—322.
- ↑ 132,0 132,1 Медников, 1994, с. 368.
- ↑ Beauchamp G. K. Sensory and receptor responses to umami: an overview of pioneering work // The American Journal of Clinical Nutrition. — 2009. — Vol. 90, № 3. — P. 723—727. — doi:10.3945/ajcn.2009.27462E. — PMID 19571221.
- ↑ 134,0 134,1 Campbell, 2011, p. 1102.
- ↑ 135,0 135,1 Зильбернагль, Деспопулос, 2013, с. 350.
- ↑ Chandrashekar J., Hoon M. A., Ryba N. J. P., Zuker C. S. The receptors and cells for mammalian taste // Nature. — 2006. — Vol. 444, № 7117. — P. 288—294. — doi:10.1038/nature05401. — PMID 17108952.
- ↑ Гистология, цитология и эмбриология, 2004, с. 303—305.
- ↑ 138,0 138,1 Лысов, Ипполитова, Максимов, Шевелёв, 2012, с. 107—109.
- ↑ Halata Z., Grim M., Bauman K. I. Friedrich Sigmund Merkel and his “Merkel cell”, morphology, development, and physiology: Review and new results // The Anatomical Record. Part A. — 2003. — Vol. 271, № 1. — P. 225—239. — doi:10.1002/ar.a.10029. — PMID 12552639.
- ↑ Halata Z., Baumann K. I., Grim M. . Merkel Nerve Endings Functioning as Mechanoreceptors in Vertebrates // The Merkel Cell: Structure — Development — Function — Cancerogenesis / Ed. by K. I. Baumann, Z. Halata, I. Moll. — Berlin, Heidelberg: Springer Verlag, 2003. — xiv + 248 p. — ISBN 978-3-642-05574-4. — P. 307—322.
- ↑ Paus R., Cotsarelis G. The biology of hair follicles // The New England Journal of Medicine. — 1999. — Vol. 341, № 7. — P. 491—497. — doi:10.1056/NEJM199908123410706. — PMID 10441606.
- ↑ Campbell, 2011, p. 1089.
- ↑ Pettigrew J. D. Electroreception in monotremes // The Journal of Experimental Biology. — 1999. — Vol. 202, № 10. — P. 1447—1454. — PMID 10210685.
- ↑ Czech-Damal N. U., Liebschner A., Miersch L. e. a. Electroreception in the Guiana dolphin (Sotalia guianensis) // Proceedings. Biological sciences / The Royal Society. — 2012. — Vol. 279, № 1729. — P. 663—668. — doi:10.1098/rspb.2011.1127. — PMID 21795271.
- ↑ Baker R. R., Mather J. G., Kennaugh J. H. Magnetic bones in human sinuses // Nature. — 1983. — Vol. 301, № 5895. — P. 79—80. — doi:10.1038/301078a0. — PMID 6823284.
- ↑ Burda H., Begall S., Červený J. e. a. Extremely low-frequency electromagnetic fields disrupt magnetic alignment of ruminants // Proc. Nat. Acad. Sci. USA. — 2009. — Vol. 106, № 14. — P. 5708–5713. — doi:10.1073/pnas.0811194106..
- ↑ Гистология, цитология и эмбриология, 2004, с. 621—623.
- ↑ Константинов, Шаталова, 2004, с. 380—381, 504—505.
- ↑ 149,0 149,1 149,2 Константинов, Наумов, Шаталова, 2012, с. 327—328.
- ↑ Гистология, цитология и эмбриология, 2004, с. 627, 630, 636.
- ↑ Дзержинский, 2005, с. 227.
- ↑ Дзержинский, 2005, с. 226—227.
- ↑ Медников, 1994, с. 361.
- ↑ 154,0 154,1 154,2 Константинов, Шаталова, 2004, с. 381.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 328.
- ↑ 156,0 156,1 Жизнь животных, т. 6, 1971, с. 12.
- ↑ Campbell, 2011, p. 925—926.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 78—80, 263—265, 401—403.
- ↑ Константинов, Шаталова, 2004, с. 203—204, 362, 382.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 81, 400—401.
- ↑ Гистология, цитология и эмбриология, 2004, с. 420—421.
- ↑ Ткаченко, 2009, с. 174—175.
- ↑ 163,0 163,1 163,2 Константинов, Шаталова, 2004, с. 383.
- ↑ Vaughan, Ryan, Czaplewski, 2011, p. 24.
- ↑ Наумов, Карташёв, 1979, с. 226.
- ↑ Медников, 1994, с. 356—357.
- ↑ 167,0 167,1 Барабаш-Никифоров, Формозов, 1963, с. 79—81.
- ↑ 168,0 168,1 Медников, 1994, с. 359.
- ↑ 169,0 169,1 169,2 Наумов, Карташёв, 1979, с. 224—225.
- ↑ Shostak S. Lecture 3. Connective Tissues Архивная копия от 21 октября 2018 на Wayback Machine
- ↑ Schmaier A. A., Stalker T. J., Runge J. J. e. a. Occlusive thrombi arise in mammals but not birds in response to arterial injury: evolutionary insight into human cardiovascular disease // Blood. — 2011. — Vol. 118, № 13. — P. 3661—3669. — doi:10.1182/blood-2011-02-338244. — PMID 21816834.
- ↑ Строгая Т. В., Евтухова Л. А. . Гистология. — Гомель: ГГУ им. Ф. Скорины, 2010. — 80 с. — ISBN 978-985-439-458-9. Архивированная копия (недоступная ссылка). Дата обращения: 2 мая 2020. Архивировано 13 апреля 2014 года. — С. 34-35.
- ↑ Гистология, цитология и эмбриология, 2004, с. 166—167, 176, 185—189.
- ↑ 174,0 174,1 Дзержинский, Васильев, Малахов, 2014, с. 403.
- ↑ Campbell, 2011, p. 911.
- ↑ Windberger U., Bartholovitsch A., Plasenzotti R. e. a. Whole blood viscosity, plasma viscosity and erythrocyte aggregation in nine mammalian species: Reference values and comparison of data // Experimental Physiology. — 2003. — Vol. 88, № 3. — P. 431—440. — PMID 12719768.
- ↑ Baskurt O. K., Marshall-Gradisnik S., Pyne M. e. a. Assessment of the hemorheological profile of koala and echidna // Zoology (Jena). — 2010. — Vol. 113, № 2. — P. 110—117. — doi:10.1016/j.zool.2009.07.003. — PMID 20138490.
- ↑ Wickham L. L., Costa D. P., Elsner R. Journal of Zoology Blood rheology of captive and free-ranging northern elephant seals and sea otters. — 1990. — Vol. 68, № 2. — P. 375–380. — doi:10.1139/z90-053..
- ↑ Castellini M. A., Baskurt O., Castellini J. M., Meiselman H. J. Blood rheology in marine mammals // Frontiers in Physiology. — 2010. — Vol. 1. — P. 146-1—146-8. — doi:10.3389/fphys.2010.00146. — PMID 21423386.
- ↑ Гистология, цитология и эмбриология, 2004, с. 166, 190.
- ↑ National Cancer Institute: The Immune System (недоступная ссылка). Дата обращения: 25 июня 2014. Архивировано 6 июля 2014 года.
- ↑ 182,0 182,1 Зильбернагль, Деспопулос, 2013, с. 100.
- ↑ Campbell, 2011, p. 930.
- ↑ Ellis S. A. Immunology: Comparative Immunology of Mammals // Wiley Online Library. — 2012. — doi:10.1002/9780470015902.a0001284.pub3.
- ↑ 185,0 185,1 Бурместер, Пецутто, 2014, с. 11.
- ↑ Галактионов, 2004, с. 10.
- ↑ Галактионов, 2004, с. 10—11.
- ↑ Галактионов, 2004, с. 13.
- ↑ Зильбернагль, Деспопулос, 2013, с. 102.
- ↑ Бурместер, Пецутто, 2014, с. 1.
- ↑ Campbell, 2011, p. 937.
- ↑ Campbell, 2011, p. 942.
- ↑ Зильбернагль, Деспопулос, 2013, с. 102—104.
- ↑ Зильбернагль, Деспопулос, 2013, с. 104.
- ↑ Campbell, 2011, p. 943.
- ↑ Ткаченко, 2009, с. 162—163.
- ↑ Campbell, 2011, p. 944.
- ↑ 198,0 198,1 Бурместер, Пецутто, 2014, p. 14.
- ↑ Taylor F. A. Animals Have Allergies Too! // The Journal of NAET® Energetics and Complementary Medicine. — 2007. — Vol. 3, № 4. — P. 849–854. Архивировано 14 июля 2014 года.
- ↑ Зильбернагль, Деспопулос, 2013, с. 106.
- ↑ Williams Z. Inducing tolerance to pregnancy // The New England Journal of Medicine. — 2012. — Vol. 367, № 12. — P. 1159—1161. — doi:10.1056/NEJMcibr1207279. — PMID 22992082.
- ↑ 202,0 202,1 Clark D. A., Chaput A., Tutton D. Active suppression of host-vs-graft reaction in pregnant mice. VII. Spontaneous abortion of allogeneic CBA/J x DBA/2 fetuses in the uterus of CBA/J mice correlates with deficient non-T suppressor cell activity // Journal of Immunology (Baltimore). — 1986. — Vol. 136, № 5. — P. 1668—1675. — PMID 2936806.
- ↑ Moffett A., Loke C. Immunology of placentation in eutherian mammals // Nature Reviews. Immunology. — 2006. — Vol. 6, № 8. — P. 584—594. — doi:10.1038/nri1897. — PMID 16868549.
- ↑ Clark G. F., Dell A., Morris H. R. e. a. The species recognition system: a new corollary for the human fetoembryonic defense system hypothesis // Cells, Tissues, Organs. — 2001. — Vol. 168, № 12. — P. 113—121. — PMID 11114593.
- ↑ Robertson S. A., Bromfield J. J., Tremellen K. P. Seminal ‘priming’ for protection from pre-eclampsia-a unifying hypothesis // Journal of Reproductive Immunology. — 2003. — Vol. 59, № 2. — P. 253—265. — doi:10.1016/S0165-0378(03)00052-4. — PMID 12896827.
- ↑ Daly T. J., Buffenstein R. Skin morphology and its role in thermoregulation in mole-rats, Heterocephalus glaber and Cryptomys hottentotus (англ.) // Journal of Anatomy. — 1998. — Vol. 193, № 4. — P. 495–502. — doi:10.1046/j.1469-7580.1998.19340495.x..
- ↑ 207,0 207,1 Дзержинский, Васильев, Малахов, 2014, с. 369, 399.
- ↑ Romanovsky A. A. Thermoregulation: some concepts have changed. Functional architecture of the thermoregulatory system // American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. — 2007. — Vol. 292, № 1. — P. 37—46. — doi:10.1152/ajpregu.00668.2006. — PMID 17008453.
- ↑ Terrien J., Perret M., Aujard F. Behavioral thermoregulation in mammals: a review // Frontiers in Bioscience (Landmark edition). — 2011. — Vol. 16. — P. 1428—1444. — PMID 21196240.
- ↑ Константинов, Шаталова, 2004, с. 385—387.
- ↑ Лютинский С. И. . Патологическая физиология животных. — М.: ГЭОТАР-Медиа, 2011. — 559 с. — ISBN 978-5-9704-1908-3. — С. 202—203.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 269, 379.
- ↑ Константинов, Шаталова, 2004, с. 385.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 310, 343.
- ↑ Разнообразие млекопитающих, ч. I, 2004, с. 192.
- ↑ Жизнь животных, т. 6, 1971, с. 102.
- ↑ Константинов, Шаталова, 2004, с. 471.
- ↑ Барабаш-Никифоров, Формозов, 1963, с. 66.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 322.
- ↑ Наумов, Карташёв, 1979, с. 215.
- ↑ Барабаш-Никифоров, Формозов, 1963, с. 66—67.
- ↑ Медников, 1994, с. 351—354.
- ↑ Жизнь животных, т. 6, 1971, с. 60, 132, 152.
- ↑ Константинов, Шаталова, 2004, с. 375.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 323.
- ↑ Кэрролл, т. 2, 1993, с. 231.
- ↑ Жизнь животных, т. 6, 1971, с. 49, 322.
- ↑ Жизнь животных, т. 6, 1971, с. 215, 304, 358, 580.
- ↑ Медников, 1994, с. 351, 401.
- ↑ Жизнь животных, т. 6, 1971, с. 121, 276.
- ↑ Диастема // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- ↑ 232,0 232,1 232,2 Ткаченко, 2009.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 322—323.
- ↑ 234,0 234,1 Медников, 1994, с. 354.
- ↑ Барабаш-Никифоров, Формозов, 1963, с. 74.
- ↑ Ligabue-Braun R., Verli H., Carlini C. R. Venomous mammals: a review // Toxicon. — 2012. — Vol. 59, № 7-8. — P. 680—695. — doi:10.1016/j.toxicon.2012.02.012. — PMID 22410495.
- ↑ Наумов, Карташёв, 1979, с. 217.
- ↑ 238,0 238,1 238,2 Наумов, Карташёв, 1979, с. 217—219.
- ↑ Жизнь животных, т. 6, 1971, с. 132, 410.
- ↑ Зильбернагль, Деспопулос, 2013, с. 250.
- ↑ Ткаченко, 2009, с. 252—253.
- ↑ Кэрролл, т. 3, 1993, с. 85.
- ↑ Жизнь животных, т. 6, 1971, с. 127, 406, 433, 438, 445, 448.
- ↑ Наумов, 1982, с. 304.
- ↑ Жизнь животных, т. 6, 1971, с. 11, 130.
- ↑ Жизнь животных, т. 6, 1971, с. 11.
- ↑ Медников, 1994, с. 353—355.
- ↑ 248,0 248,1 248,2 248,3 Наумов, Карташёв, 1979, с. 219—220.
- ↑ Dennis E. A. Diversity of group types, regulation, and function of phospholipase A2 // The Journal of Biological Chemistry. — 1994. — Vol. 269, № 18. — P. 13057—13060. — PMID 8175726.
- ↑ Ткаченко, 2009, с. 257—258.
- ↑ Зильбернагль, Деспопулос, 2013, с. 254.
- ↑ Желчь — статья из Научно-технического словаря
- ↑ Ткаченко, 2009, с. 259.
- ↑ Зильбернагль, Деспопулос, 2013, с. 256.
- ↑ Зильбернагль, Деспопулос, 2013, с. 262.
- ↑ Barret K. E. . Gastrointestinal Physiology. — New York: McGraw-Hill, 2005. — xiv + 295 p. — (Lange Physiology Series). — ISBN 0-07-142310-9. — P. 156.
- ↑ Romer A. S., Parsons T. S. . The Vertebrate Body. 5th ed. — Philadelphia: W. B. Saunders, 1977. — viii + 624 p. — ISBN 0-03-910284-X. — P. 353—354.
- ↑ Жизнь животных, т. 6, 1971, с. 58.
- ↑ Разнообразие млекопитающих, ч. II, 2004, с. 379.
- ↑ Барабаш-Никифоров, Формозов, 1963, с. 133.
- ↑ Hirakawa H. Coprophagy in leporids and other mammalian herbivores // Mammal Review. — 2001. — Vol. 31, № 1. — P. 61–80. — doi:10.1046/j.1365-2907.2001.00079.x..
- ↑ 262,0 262,1 Медников, 1994, с. 355.
- ↑ 263,0 263,1 Наумов, 1982, с. 305.
- ↑ Наумов, Карташёв, 1979, с. 220.
- ↑ Campbell, 2011, p. 962.
- ↑ Медников, 1994, с. 362.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 338.
- ↑ Ткаченко, 2009, с. 305.
- ↑ Гистология, цитология и эмбриология, 2004, с. 692—693.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 337.
- ↑ 271,0 271,1 Гистология, цитология и эмбриология, 2004, с. 678—679.
- ↑ 272,0 272,1 272,2 Ткаченко, 2009, с. 305—310.
- ↑ 273,0 273,1 273,2 Campbell, 2011, p. 963—965.
- ↑ Campbell, 2011, p. 969—971.
- ↑ Ткаченко, 2009, с. 306—307.
- ↑ Campbell, 2011, p. 967.
- ↑ Тейлор Д., Грин Н., Стаут У. . Биология: в 3-х тт. Т. 3. — М.: Мир, 2010. — 451 с. — ISBN 978-5-03-003828-5. — С. 7.
- ↑ Наумов, Карташёв, 1979, с. 235.
- ↑ Campbell, 2011, p. 988.
- ↑ Ткаченко, 2009, с. 81—82.
- ↑ Ткаченко, 2009, с. 103, 112.
- ↑ 282,0 282,1 Campbell, 2011, p. 986.
- ↑ Ткаченко, 2009, с. 121.
- ↑ Ткаченко, 2009, с. 123.
- ↑ Ткаченко, 2009, с. 113—115.
- ↑ Ткаченко, 2009, с. 126—127.
- ↑ Elayat A. A., el-Naggar M. M., Tahir M. An immunocytochemical and morphometric study of the rat pancreatic islets // Journal of Anatomy. — 1995. — Vol. 186 (Pt. 3). — P. 629—637. — PMID 7559135.
- ↑ Hormones of the Liver (недоступная ссылка). Дата обращения: 11 августа 2014. Архивировано 6 июля 2014 года.
- ↑ Ткаченко, 2009, с. 127—128.
- ↑ Campbell, 2011, p. 976.
- ↑ 291,0 291,1 291,2 291,3 Ткаченко, 2009, с. 133.
- ↑ Ткаченко, 2009, с. 264.
- ↑ Зильбернагль, Деспопулос, 2013, с. 312.
- ↑ Humbio: Гормоны тимуса (недоступная ссылка). Архивировано 7 марта 2014 года.
- ↑ 295,0 295,1 Ткаченко, 2009, с. 129.
- ↑ MedicineNet: erythropoietin. Дата обращения: 21 июня 2014. Архивировано 9 августа 2012 года.
- ↑ Ткаченко, 2009, с. 132.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 406—407.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 340—346.
- ↑ Константинов, Шаталова, 2004, с. 394.
- ↑ Барабаш-Никифоров, Формозов, 1963, с. 86—88, 267.
- ↑ Яичниковый цикл — статья из Большого медицинского словаря
- ↑ 303,0 303,1 303,2 303,3 Campbell, 2011, p. 1010.
- ↑ Ткаченко, 2009, с. 469—470.
- ↑ Ткаченко, 2009, с. 470—471.
- ↑ 306,0 306,1 Vaughan, Ryan, Czaplewski, 2011, p. 392.
- ↑ Vaughan, Ryan, Czaplewski, 2011, p. 392—395.
- ↑ Estrous Cycle Learning Module. Дата обращения: 23 июня 2014. Архивировано 1 марта 2014 года.
- ↑ Estrous cycle (недоступная ссылка). Дата обращения: 23 июня 2014. Архивировано 5 марта 2016 года.
- ↑ Vaughan, Ryan, Czaplewski, 2011, p. 392—394.
- ↑ The Laboratory Rat. 2nd ed / Ed. by M. A. Suckow, S. H. Weisbroth, C. L. Franklin. — London: Elsevier Academic Press, 2006. — xvi + 912 p. — ISBN 978-0-12-074903-4. — P. 149.
- ↑ Engelking L. R. . Review of Veterinary Physiology. — Jackson: Teton NewMedia, 2002. — 657 p. — ISBN 1-893441-69-5. — P. 436.
- ↑ Гродницкий Д. Л. Концепция зародышевой плазмы Августа Вейсмана как основная причина неадекватности неодарвинизма // Журнал общей биологии. — 2000. — Т. 61, № 4. — С. 371—380.
- ↑ Warren W. C., Hillier L. W., Marshall Graves J. A. e. a. Genome analysis of the Platypus reveals unique signatures of evolution // Nature. — 2008. — Vol. 453, № 7192. — P. 175—183. — doi:10.1038/nature06936. — PMID 18464734.
- ↑ Marshall Graves J. A. The rise and fall of SRY // Trends in Genetics. — 2002. — Vol. 18, № 5. — P. 259—264. — doi:10.1016/S0168-9525(02)02666-5. — PMID 12047951.
- ↑ Kuroiwa A., Handa S., Nishiyama C. e. a. Additional copies of CBX2 in the genomes of males of mammals lacking SRY, the Amami spiny rat (Tokudaia osimensis) and the Tokunoshima spiny rat (Tokudaia tokunoshimensis) // Chromosome Research. — 2011. — Vol. 19, № 5. — P. 635—644. — doi:10.1007/s10577-011-9223-6. — PMID 21656076.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 408.
- ↑ Vaughan, Ryan, Czaplewski, 2011, p. 25, 389.
- ↑ Разнообразие млекопитающих, ч. I, 2004, с. 90—91, 95.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 408—410.
- ↑ 321,0 321,1 Кэрролл, т. 3, 1993, с. 5—6, 241.
- ↑ Разнообразие млекопитающих, ч. I, 2004, с. 109, 114, 136, 142, 176.
- ↑ Chelini M. O., Souza N. L., Rocha A. M. e. a. Quantification of fecal estradiol and progesterone metabolites in Syrian hamsters (Mesocricetus auratus) // Brazilian Journal of Medical and Biological Research. — 2005. — Vol. 38, № 11. — P. 1711–1717.
- ↑ Hautier L., Stansfield F. J., Allen W. R., Asher R. J. Skeletal development in the African elephant and ossification timing in placental mammals // Proceedings. Biological Sciences / The Royal Society. — 2012. — Vol. 279, № 1736. — P. 2188—2195. — doi:10.1098/rspb.2011.2481. — PMID 22298853.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 410.
- ↑ 326,0 326,1 Average Animal Gestation Periods and Incubation Times
- ↑ Разнообразие млекопитающих, ч. II, 2004, с. 390, 398, 412, 571.
- ↑ Разнообразие млекопитающих, ч. III, 2004, с. 815.
- ↑ Yamaguchi N., Dugdale H. L., Macdonald D. W. Female receptivity, embryonic diapause, and superfetation in the European badger (Meles meles): implications for the reproductive tactics of males and females // The Quarterly Review of Biology. — 2006. — Vol. 81, № 1. — P. 33—48. — PMID 16602273.
- ↑ Larivière S., Walton L. R. Lontra canadensis (англ.) // Mammalian Species. — American Society of Mammalogists, 1998. — No. 587. — P. 1–8.
- ↑ Wild Mammals of North America: Biology, Management, and Conservation. 2nd ed / Ed. by G. A. Feldhamer, B. C. Thompson, J. A. Chapman. — Baltimore: The Johns Hopkins University Press, 2003. — xiii + 1216 p. — ISBN 0-8018-7416-5. — P. 664, 685, 714.
- ↑ Разнообразие млекопитающих, ч. III, 2004, с. 906.
- ↑ Vaughan, Ryan, Czaplewski, 2011, p. 311.
- ↑ Sears R., Perrin W. F. . Blue Whale. Balaenoptera musculus // Encyclopedia of Marine Mammals. 2nd ed / Ed. by W. F. Perrin, B. Würsig, J. G. M. Thewissen. — New York: Academic Press, 2005. — xxxiv + 1316 p. — ISBN 978-0-12-373553-9. — P. 120—125.
- ↑ Barton N. H., Briggs D. E. G., Eisen J. A. . Evolution. — Cold Spring Harbour: Cold Spring Harbor Laboratory Press, U.S., 2007. — 833 p. — ISBN 978-0-87-969684-9. — P. 581—583.
- ↑ McPherson F. J., Chenoweth P. J. Mammalian sexual dimorphism // Animal Reproduction Science. — 2012. — Vol. 131, № 3–4. — P. 109–122. — doi:10.1016/j.anireprosci.2012.02.007.
Литература
На русском языке
- Барабаш-Никифоров И. И., Формозов А. Н. . Териология. — М.: Высшая школа, 1963. — 396 с.
- Бурместер Г.-Р., Пецутто А. Наглядная иммунология = Color Atlas of Immunology : [пер. с англ.] / под ред. профессора, доктора биол. наук Козлова Л. В.. — 3-е изд. — М. : БИНОМ. Лаборатория знаний, 2014. — 320 с. : ил. — (Наглядная медицина). — ББК 52.54. — УДК 612.017(G). — ISBN 978-5-9963-1731-8.
- Галактионов В. Г. . Иммунология. — М.: Издат. центр «Академия», 2004. — 528 с. — ISBN 5-7695-1260-1.
- Гистология, цитология и эмбриология. 6-е изд / Под ред. Ю. И. Афанасьева, С. Л. Кузнецова, H. А. Юриной. — М.: Медицина, 2004. — 768 с. — ISBN 5-225-04858-7.
- Дзержинский Ф. Я. . Сравнительная анатомия позвоночных животных. — М.: Аспект пресс, 2005. — 304 с. — ISBN 5-7567-0360-8.
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. . Зоология позвоночных. 2-е изд. — М.: Издат. центр «Академия», 2014. — 464 с. — ISBN 978-5-4468-0459-7.
- Жизнь животных. Энциклопедия в 6 тт. Т. 6: Млекопитающие / Общ. ред. Л. А. Зенкевича. — М.: Просвещение, 1971. — 628 с.
- Зильбернагль С., Деспопулос А. . Наглядная физиология. — М.: БИНОМ. Лаборатория знаний, 2013. — 408 с. — ISBN 978-5-94774-385-2.
- Константинов В. М., Наумов С. П., Шаталова С. П. . Зоология позвоночных. 7-е изд. — М.: Издат. центр «Академия», 2012. — 448 с. — ISBN 978-5-7695-9293-5.
- Константинов В. М., Шаталова С. П. . Зоология позвоночных. — М.: Гуманитарный издательский центр ВЛАДОС, 2004. — 527 с. — ISBN 5-691-01293-2.
- Кэрролл Р. . Палеонтология и эволюция позвоночных: В 3-х тт. Т. 2. — М.: Мир, 1993. — 283 с. — ISBN 5-03-001819-0.
- Кэрролл Р. . Палеонтология и эволюция позвоночных: В 3-х тт. Т. 3. — М.: Мир, 1993. — 312 с. — ISBN 5-03-001819-0.
- Лысов В. Ф., Ипполитова Т. В., Максимов В. И., Шевелёв Н. С. . Физиология и этология животных. 2-е изд. — М.: КолосС, 2012. — 605 с. — ISBN 978-5-9532-0826-0.
- Медников Б. М. . Биология: формы и уровни жизни. — М.: Просвещение, 1994. — 415 с. — ISBN 5-09-004384-1.
- Наумов Н. П., Карташёв Н. Н. . Зоология позвоночных. Ч. 2. Пресмыкающиеся, птицы, млекопитающие. — М.: Высшая школа, 1979. — 272 с.
- Наумов С. П. . Зоология позвоночных. — М.: Просвещение, 1982. — 464 с.
- Разнообразие млекопитающих / О. Л. Россолимо, И. Я. Павлинов, С. В. Крускоп, А. А. Лисовский, Н. Н. Спасская, А. В. Борисенко, А. А. Панютина. — М. : Издательство КМК, 2004. — Ч. I. — 366 с. — (Разнообразие животных). — ISBN 5-87317-098-3.
- Разнообразие млекопитающих / О. Л. Россолимо, И. Я. Павлинов, С. В. Крускоп, А. А. Лисовский, Н. Н. Спасская, А. В. Борисенко, А. А. Панютина. — М. : Издательство КМК, 2004. — Ч. II. — 218 с. — (Разнообразие животных). — ISBN 5-87317-098-3.
- Разнообразие млекопитающих / О. Л. Россолимо, И. Я. Павлинов, С. В. Крускоп, А. А. Лисовский, Н. Н. Спасская, А. В. Борисенко, А. А. Панютина. — М. : Издательство КМК, 2004. — Ч. III. — 408 с. — (Разнообразие животных). — ISBN 5-87317-098-3.
- Сидоров А. В. . Физиология межклеточной коммуникации. — Минск: БГУ, 2008. — 215 с. — ISBN 978-985-485-812-8.
- Ткаченко Б. И., Брин В. Б., Захаров Ю. М., Недоспасов В. О., Пятин В. Ф. . Физиология человека. Compendium / Под ред. Б. И. Ткаченко. — М.: ГЭОТАР-Медиа, 2009. — 496 с. — ISBN 978-5-9704-0964-0.
- Флинт В. Е., Чугунов Ю. Д., Смирин В. А. . Млекопитающие СССР. 2-е изд / Отв. ред. А. Н. Формозов. — М.: Мысль, 1970. — 437 с.
На английском языке
- Campbell N. A., Reece J. B., Urry L. A. e. a. . Biology. 9th ed. — Benjamin Cummings, 2011. — 1263 p. — ISBN 978-0-321-55823-7.
- Vaughan T. A., Ryan J. M., Czaplewski N. J. . Mammalogy. 5th ed. — Sudbury, Massachusetts: Jones & Bartlett Learning, 2011. — 750 p. — ISBN 978-0-7636-6299-5.
Ссылки
- Электронная библиотека книг по млекопитающим Архивная копия от 10 декабря 2013 на Wayback Machine (Дата обращения: 16 февраля 2014)
Эта статья входит в число золотых статей пантеона энциклопедии Руниверсалис. |